

Lipid storage by adipose tissue macrophages regulates systemic glucose tolerance
- PMID: 24986598
- PMCID: PMC4137117
- DOI: 10.1152/ajpendo.00187.2014
Lipid storage by adipose tissue macrophages regulates systemic glucose tolerance
Abstract
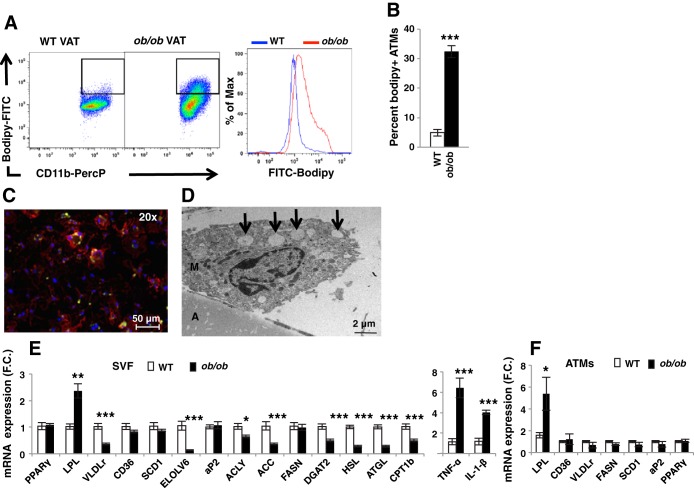
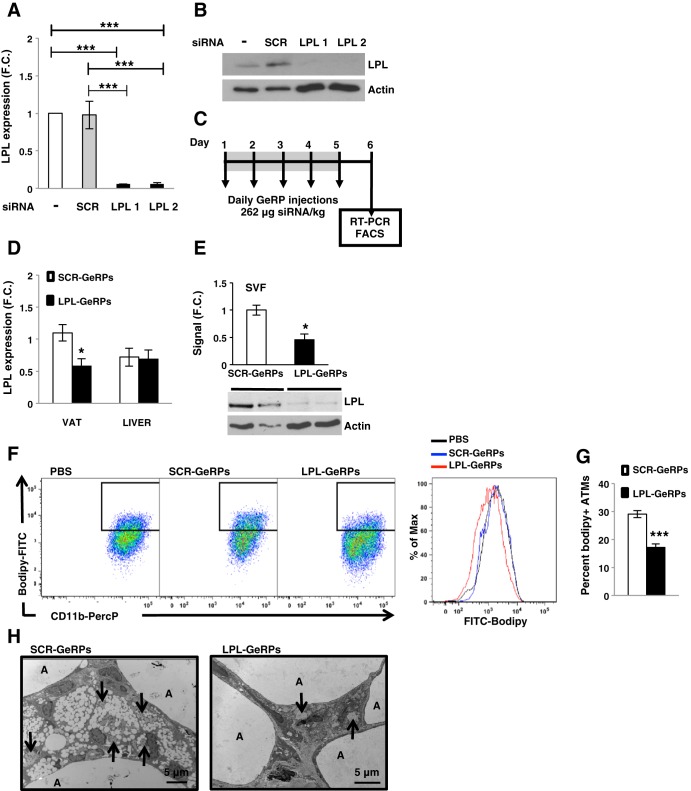
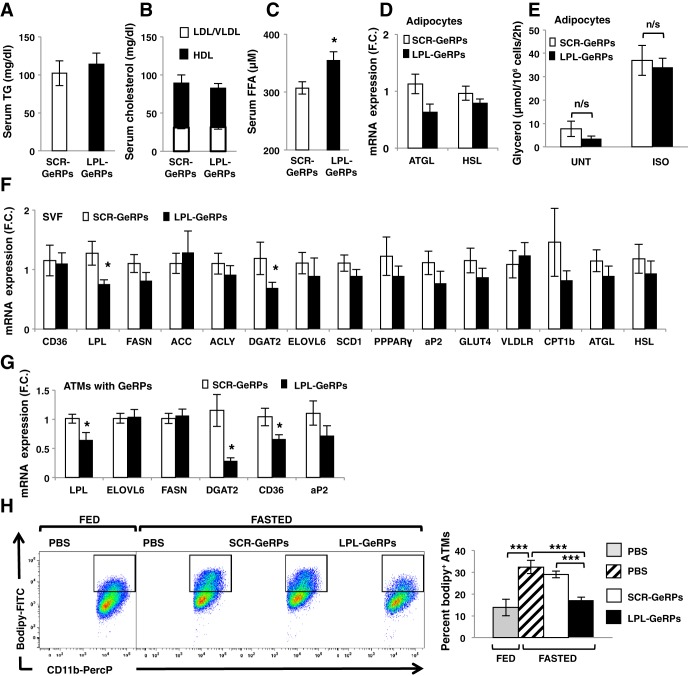
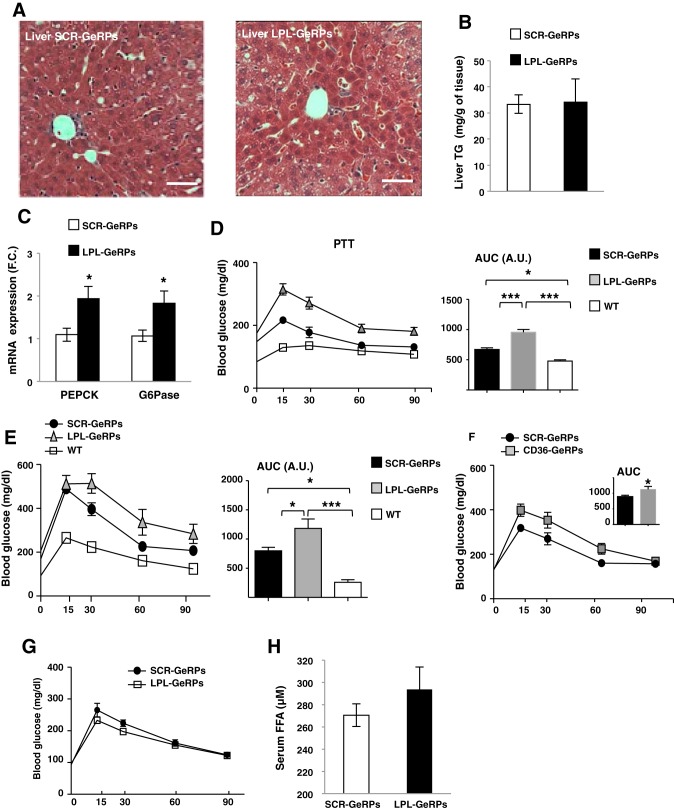
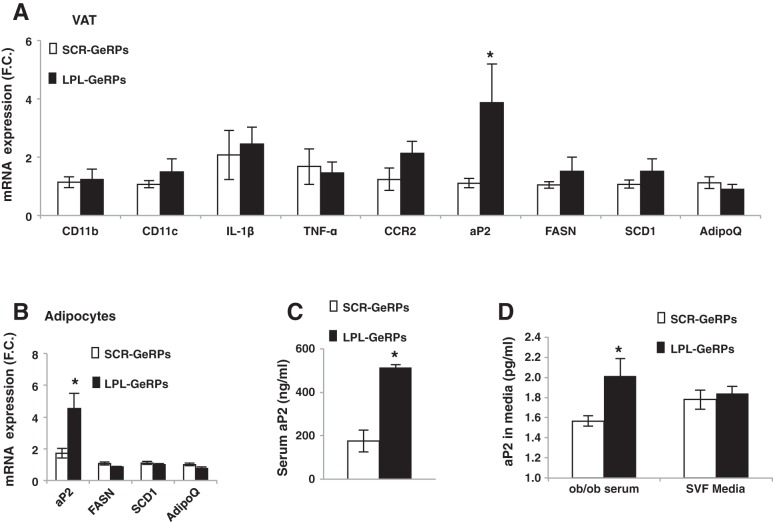
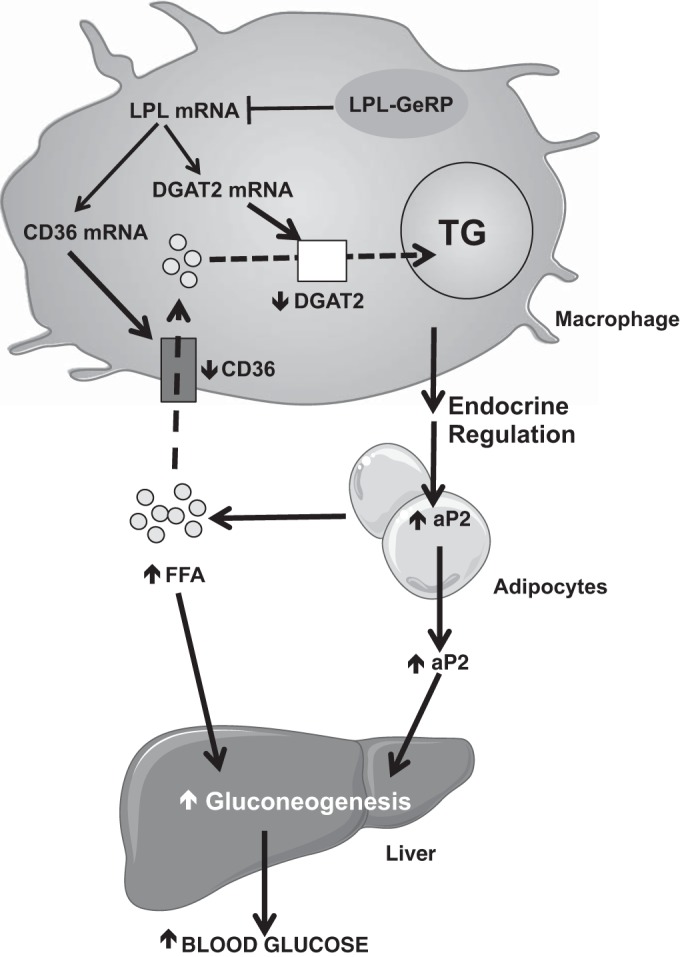
Proinflammatory pathways in adipose tissue macrophages (ATMs) can impair glucose tolerance in obesity, but ATMs may also be beneficial as repositories for excess lipid that adipocytes are unable to store. To test this hypothesis, we selectively targeted visceral ATMs in obese mice with siRNA against lipoprotein lipase (LPL), leaving macrophages within other organs unaffected. Selective silencing of ATM LPL decreased foam cell formation in visceral adipose tissue of obese mice, consistent with a reduced supply of fatty acids from VLDL hydrolysis. Unexpectedly, silencing LPL also decreased the expression of genes involved in fatty acid uptake (CD36) and esterification in ATMs. This deficit in fatty acid uptake capacity was associated with increased circulating serum free fatty acids. Importantly, ATM LPL silencing also caused a marked increase in circulating fatty acid-binding protein-4, an adipocyte-derived lipid chaperone previously reported to induce liver insulin resistance and glucose intolerance. Consistent with this concept, obese mice with LPL-depleted ATMs exhibited higher hepatic glucose production from pyruvate and glucose intolerance. Silencing CD36 in ATMs also promoted glucose intolerance. Taken together, the data indicate that LPL secreted by ATMs enhances their ability to sequester excess lipid in obese mice, promoting systemic glucose tolerance.
Keywords: adipose tissue macrophages; foam cells; insulin resistance; obesity; siRNA.
Copyright © 2014 the American Physiological Society.
Figures
References
-
- Babaev VR, Patel MB, Semenkovich CF, Fazio S, Linton MF. Macrophage lipoprotein lipase promotes foam cell formation and atherosclerosis in low density lipoprotein receptor-deficient mice. J Biol Chem 275: 26293–26299, 2000 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous