

Alzheimer's associated β-amyloid protein inhibits influenza A virus and modulates viral interactions with phagocytes
- PMID: 24988208
- PMCID: PMC4079246
- DOI: 10.1371/journal.pone.0101364
Alzheimer's associated β-amyloid protein inhibits influenza A virus and modulates viral interactions with phagocytes
Abstract
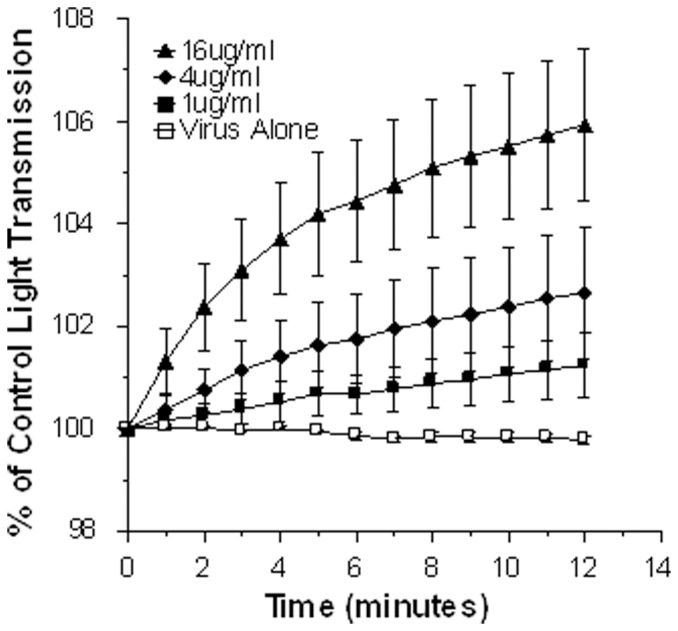
Accumulation of β-Amyloid (βA) is a key pathogenetic factor in Alzheimer's disease; however, the normal function of βA is unknown. Recent studies have shown that βA can inhibit growth of bacteria and fungi. In this paper we show that βA also inhibits replication of seasonal and pandemic strains of H3N2 and H1N1 influenza A virus (IAV) in vitro. The 42 amino acid fragment of βA (βA42) had greater activity than the 40 amino acid fragment. Direct incubation of the virus with βA42 was needed to achieve optimal inhibition. Using quantitative PCR assays βA42 was shown to reduce viral uptake by epithelial cells after 45 minutes and to reduce supernatant virus at 24 hours post infection. βA42 caused aggregation of IAV particles as detected by light transmission assays and electron and confocal microscopy. βA42 did not stimulate neutrophil H2O2 production or extracellular trap formation on its own, but it increased both responses stimulated by IAV. In addition, βA42 increased uptake of IAV by neutrophils. βA42 reduced viral protein synthesis in monocytes and reduced IAV-induced interleukin-6 production by these cells. Hence, we demonstrate for the first time that βA has antiviral activity and modulates viral interactions with phagocytes.
Conflict of interest statement
Figures








References
-
- Dahlgren KN, Manelli AM, Stine WB Jr, Baker LK, Krafft GA, et al. (2002) Oligomeric and fibrillar species of amyloid-beta peptides differentially affect neuronal viability. J Biol Chem 277: 32046–32053. - PubMed
-
- Selkoe DJ (2008) Biochemistry and molecular biology of amyloid beta-protein and the mechanism of Alzheimer's disease. Handb Clin Neurol 89: 245–260. - PubMed
-
- Papareddy P, Morgelin M, Walse B, Schmidtchen A, Malmsten M (2012) Antimicrobial activity of peptides derived from human ss-amyloid precursor protein. J Pept Sci 18: 183–191. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
