

BeMADS1 is a key to delivery MADSs into nucleus in reproductive tissues-De novo characterization of Bambusa edulis transcriptome and study of MADS genes in bamboo floral development
- PMID: 24989161
- PMCID: PMC4087239
- DOI: 10.1186/1471-2229-14-179
BeMADS1 is a key to delivery MADSs into nucleus in reproductive tissues-De novo characterization of Bambusa edulis transcriptome and study of MADS genes in bamboo floral development
Abstract
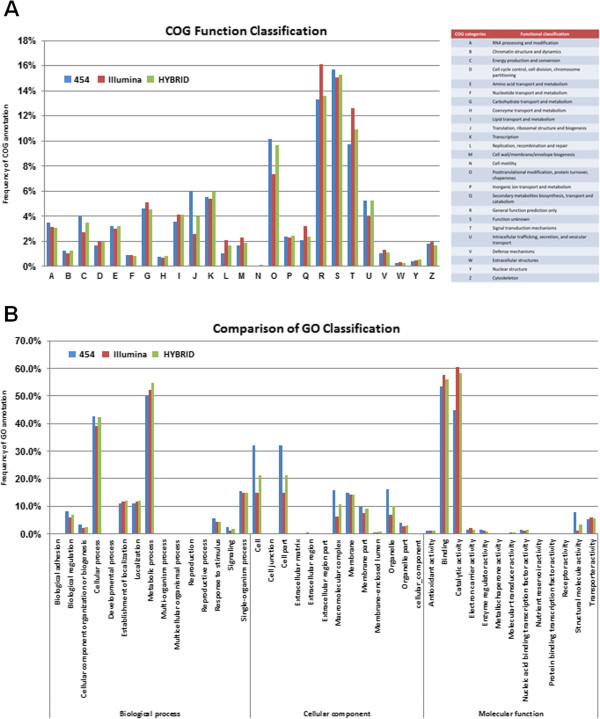
Background: The bamboo Bambusa edulis has a long juvenile phase in situ, but can be induced to flower during in vitro tissue culture, providing a readily available source of material for studies on reproductive biology and flowering. In this report, in vitro-derived reproductive and vegetative materials of B. edulis were harvested and used to generate transcriptome databases by use of two sequencing platforms: Illumina and 454. Combination of the two datasets resulted in high transcriptome quality and increased length of the sequence reads. In plants, many MADS genes control flower development, and the ABCDE model has been developed to explain how the genes function together to create the different whorls within a flower.
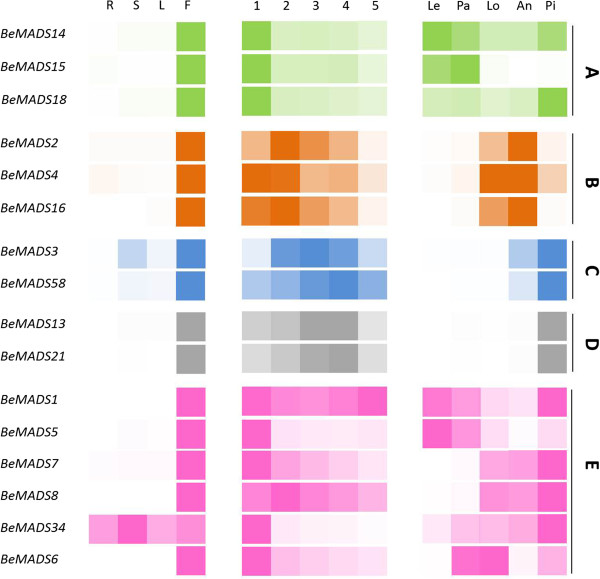
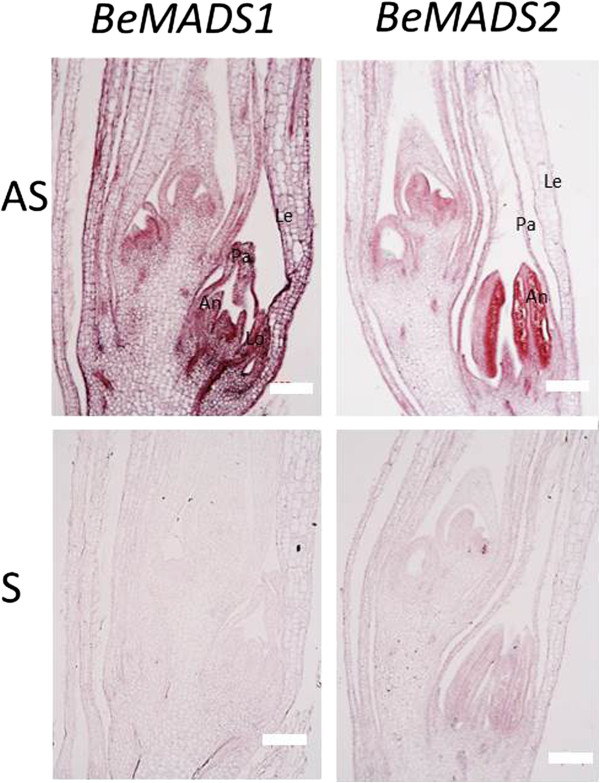
Results: As a case study, published floral development-related OsMADS proteins from rice were used to search the B. edulis transcriptome datasets, identifying 16 B. edulis MADS (BeMADS). The BeMADS gene expression levels were determined qRT-PCR and in situ hybridization. Most BeMADS genes were highly expressed in flowers, with the exception of BeMADS34. The expression patterns of these genes were most similar to the rice homologs, except BeMADS18 and BeMADS34, and were highly similar to the floral development ABCDE model in rice. Transient expression of MADS-GFP proteins showed that only BeMADS1 entered leaf nucleus. BeMADS18, BeMADS4, and BeMADS1 were located in the lemma nucleus. When co-transformed with BeMADS1, BeMADS15, 16, 13, 21, 6, and 7 translocated to nucleus in lemmas, indicating that BeMADS1 is a key factor for subcellular localization of other BeMADS.
Conclusion: Our study provides abundant B. edulis transcriptome data and offers comprehensive sequence resources. The results, molecular materials and overall strategy reported here can be used for future gene identification and for further reproductive studies in the economically important crop of bamboo.
Figures




References
-
- Li Z, Denich M. Is Shennongjia a suitable site for reintroducing giant panda: an appraisal on food supply. Environmentalist. 2004;24:165–170.
-
- Xiong W. The present and future situation of bamboo industry in Japan. Bamboo Res. 1982;1:92–93.
-
- Nadgauda RS, Parasharami VA, Mascarenhas AF. Precocious flowering and seeding behavior in tissue cultured bamboos. Nature. 1990;344:335–336.
-
- Lin CS, Chang WC. Micropropagation of Bambusa edulis through nodal explants of field-grown culms and flowering of regenerated plantlets. Plant Cell Rep. 1998;17:617–620. - PubMed
-
- Lin CS, Lin CC, Chang WC. In vitro flowering of Bambusa edulis and subsequent plantlet survival. Plant Cell Tiss Org. 2003;72:71–78.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
