

Epigenetic regulation of pluripotency and differentiation
- PMID: 24989490
- PMCID: PMC4229506
- DOI: 10.1161/CIRCRESAHA.115.301517
Epigenetic regulation of pluripotency and differentiation
Abstract
The precise, temporal order of gene expression during development is critical to ensure proper lineage commitment, cell fate determination, and ultimately, organogenesis. Epigenetic regulation of chromatin structure is fundamental to the activation or repression of genes during embryonic development. In recent years, there has been an explosion of research relating to various modes of epigenetic regulation, such as DNA methylation, post-translational histone tail modifications, noncoding RNA control of chromatin structure, and nucleosome remodeling. Technological advances in genome-wide epigenetic profiling and pluripotent stem cell differentiation have been primary drivers for elucidating the epigenetic control of cellular identity during development and nuclear reprogramming. Not only do epigenetic mechanisms regulate transcriptional states in a cell-type-specific manner but also they establish higher order genomic topology and nuclear architecture. Here, we review the epigenetic control of pluripotency and changes associated with pluripotent stem cell differentiation. We focus on DNA methylation, DNA demethylation, and common histone tail modifications. Finally, we briefly discuss epigenetic heterogeneity among pluripotent stem cell lines and the influence of epigenetic patterns on genome topology.
Keywords: epigenomics; methylation; stem cell, pluripotent.
© 2014 American Heart Association, Inc.
Figures
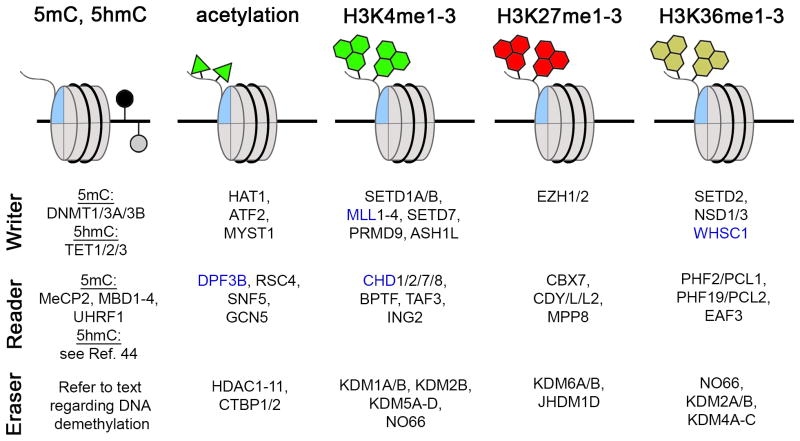
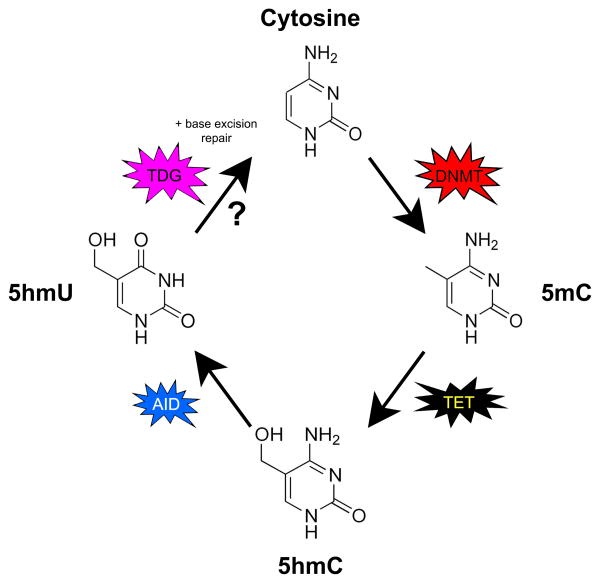
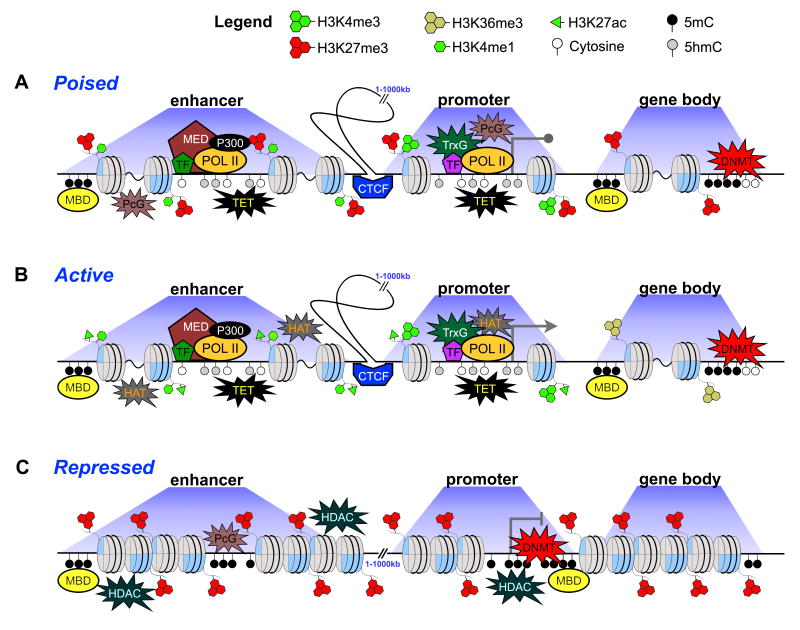
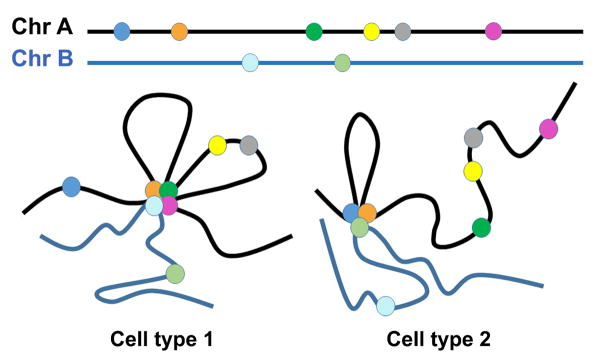
References
-
- Gaspard N, Bouschet T, Hourez R, Dimidschstein J, Naeije G, van den Ameele J, Espuny-Camacho I, Herpoel A, Passante L, Schiffmann SN, Gaillard A, Vanderhaeghen P. An intrinsic mechanism of corticogenesis from embryonic stem cells. Nature. 2008;455:351–357. - PubMed
-
- Espuny-Camacho I, Michelsen Kimmo A, Gall D, Linaro D, Hasche A, Bonnefont J, Bali C, Orduz D, Bilheu A, Herpoel A, Lambert N, Gaspard N, Péron S, Schiffmann Serge N, Giugliano M, Gaillard A, Vanderhaeghen P. Pyramidal neurons derived from human pluripotent stem cells integrate efficiently into mouse brain circuits in vivo. Neuron. 2013;77:440–456. - PubMed
-
- Eiraku M, Takata N, Ishibashi H, Kawada M, Sakakura E, Okuda S, Sekiguchi K, Adachi T, Sasai Y. Self-organizing optic-cup morphogenesis in three-dimensional culture. Nature. 2011;472:51–56. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
