

Structural dynamics of the two-component response regulator RstA in recognition of promoter DNA element
- PMID: 24990372
- PMCID: PMC4117788
- DOI: 10.1093/nar/gku572
Structural dynamics of the two-component response regulator RstA in recognition of promoter DNA element
Abstract
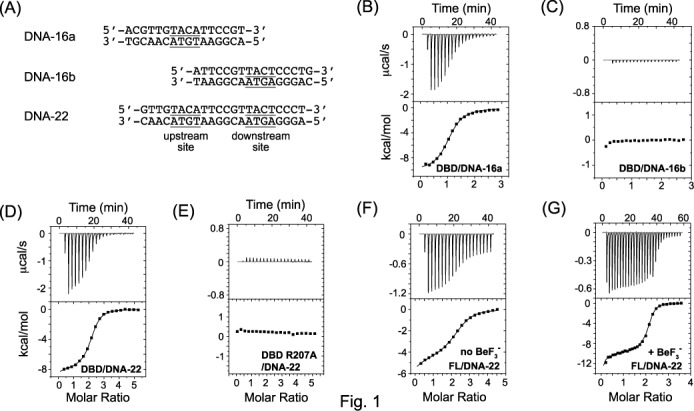
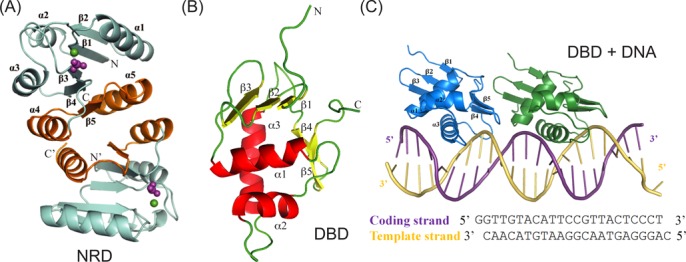
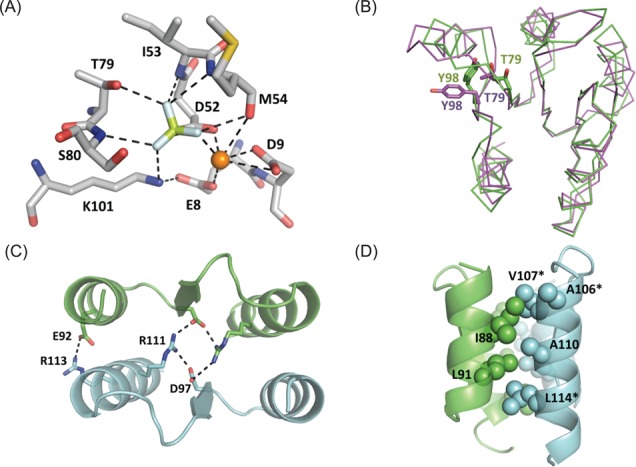
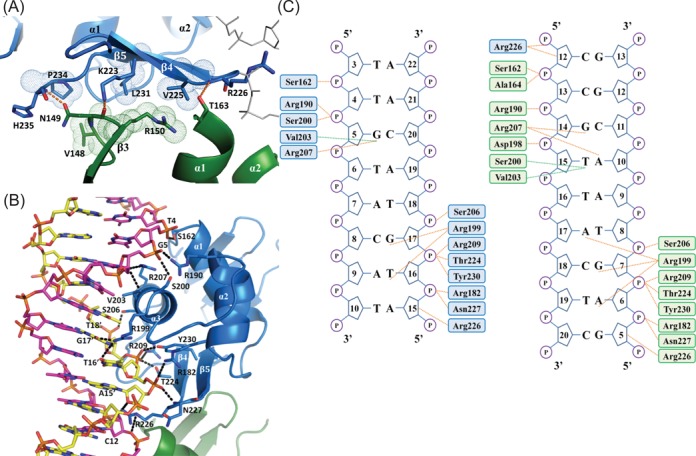
The RstA/RstB system is a bacterial two-component regulatory system consisting of the membrane sensor, RstB and its cognate response regulator (RR) RstA. The RstA of Klebsiella pneumoniae (kpRstA) consists of an N-terminal receiver domain (RD, residues 1-119) and a C-terminal DNA-binding domain (DBD, residues 130-236). Phosphorylation of kpRstA induces dimerization, which allows two kpRstA DBDs to bind to a tandem repeat, called the RstA box, and regulate the expression of downstream genes. Here we report the solution and crystal structures of the free kpRstA RD, DBD and DBD/RstA box DNA complex. The structure of the kpRstA DBD/RstA box complex suggests that the two protomers interact with the RstA box in an asymmetric fashion. Equilibrium binding studies further reveal that the two protomers within the kpRstA dimer bind to the RstA box in a sequential manner. Taken together, our results suggest a binding model where dimerization of the kpRstA RDs provides the platform to allow the first kpRstA DBD protomer to anchor protein-DNA interaction, whereas the second protomer plays a key role in ensuring correct recognition of the RstA box.
© The Author(s) 2014. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
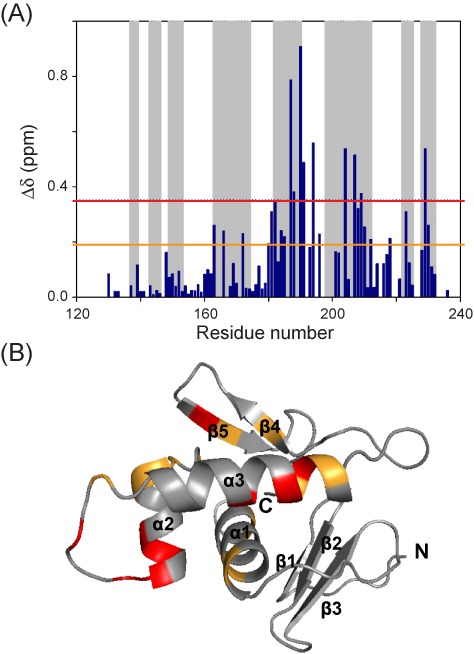
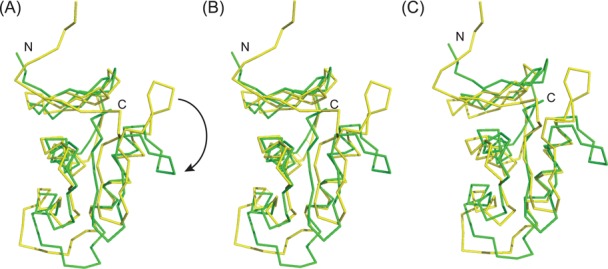
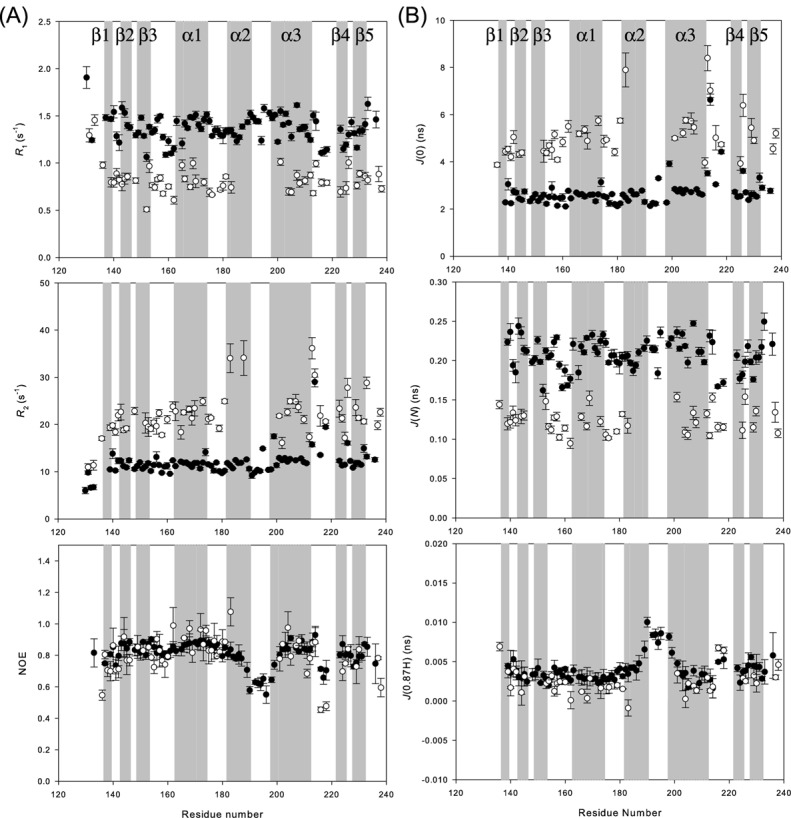
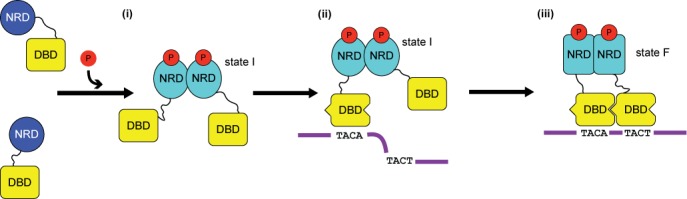
References
-
- Parkinson J.S., Kofoid E.C. Communication modules in bacterial signaling proteins. Annu. Rev. Genet. 1992;26:71–112. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
