

Endothelial cells translate pathogen signals into G-CSF-driven emergency granulopoiesis
- PMID: 24990886
- PMCID: PMC4148762
- DOI: 10.1182/blood-2014-04-570762
Endothelial cells translate pathogen signals into G-CSF-driven emergency granulopoiesis
Abstract
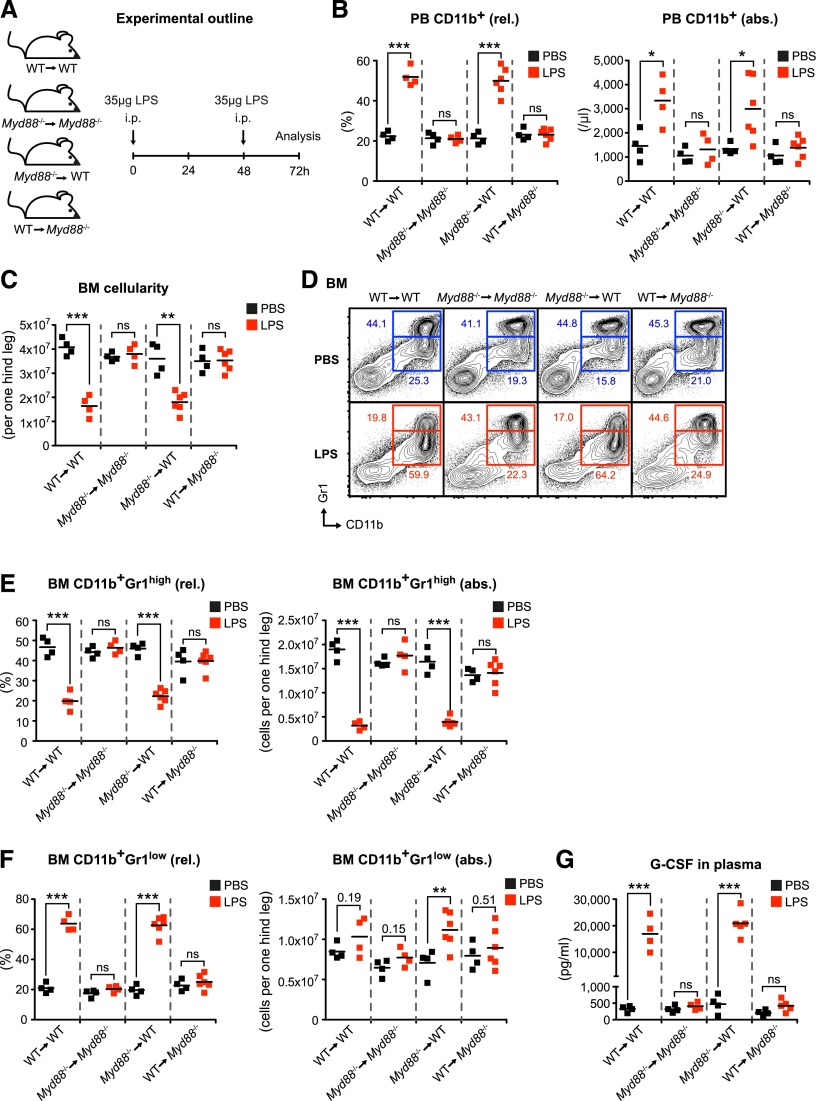
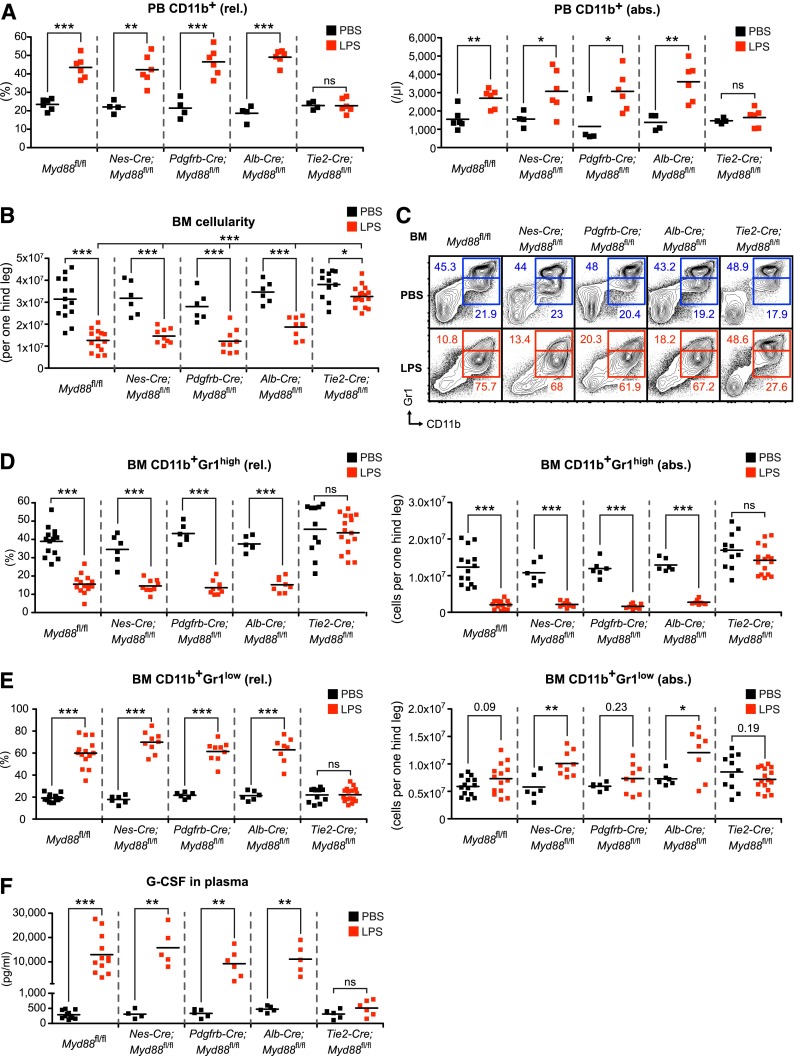
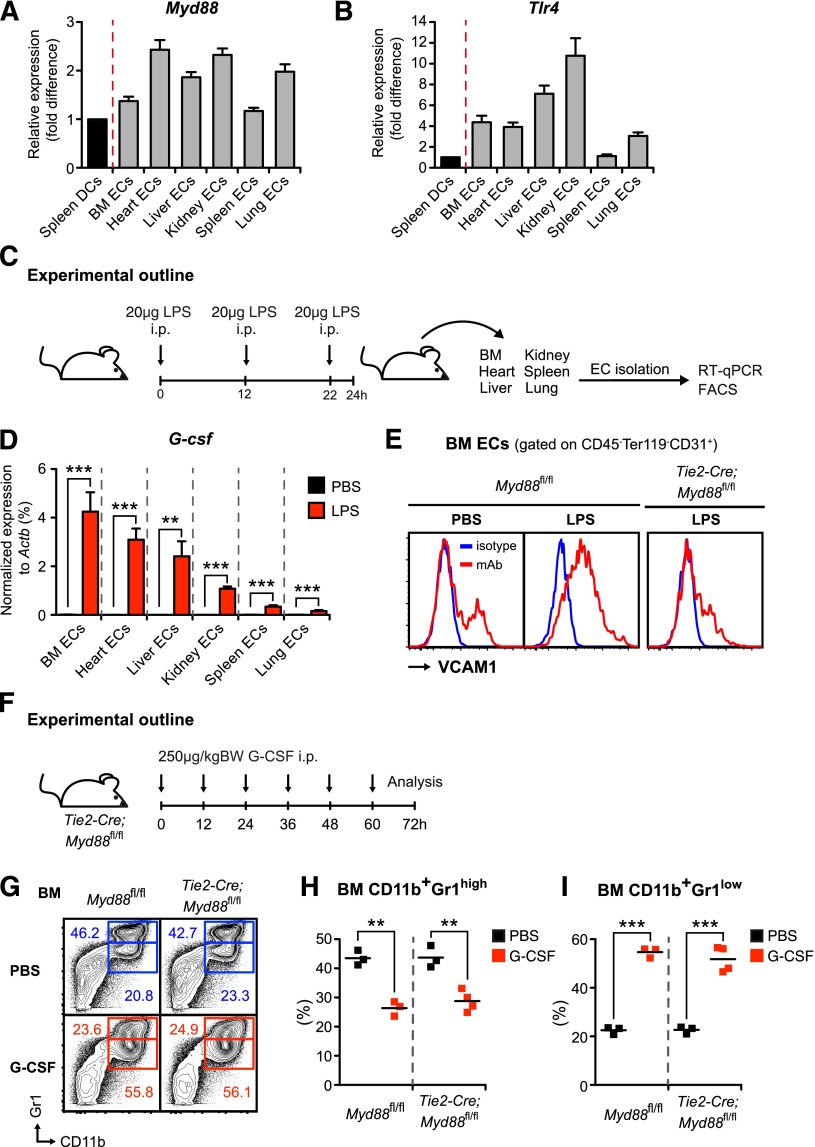
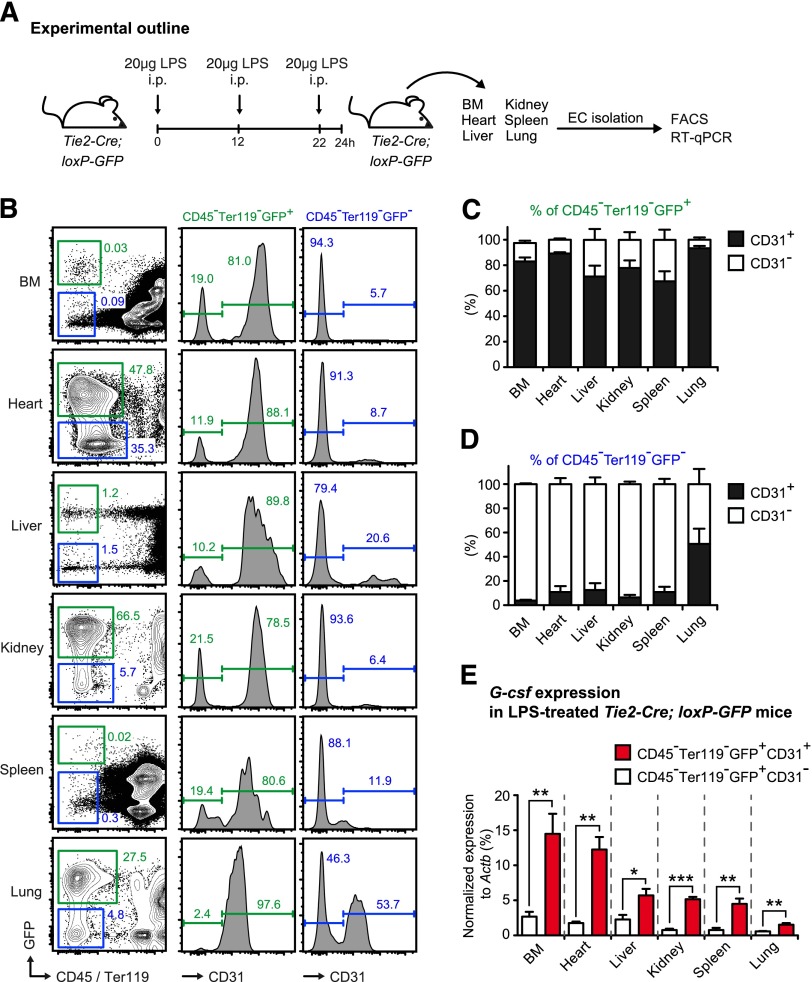
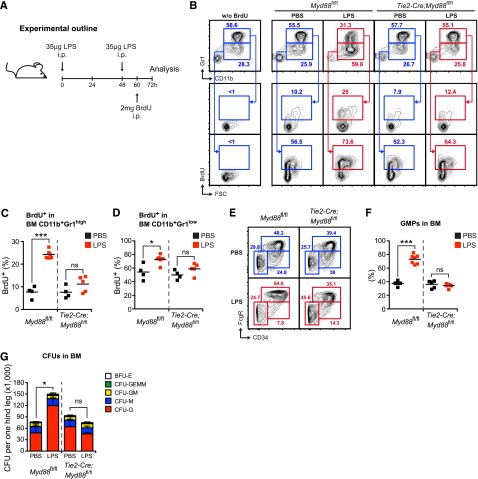
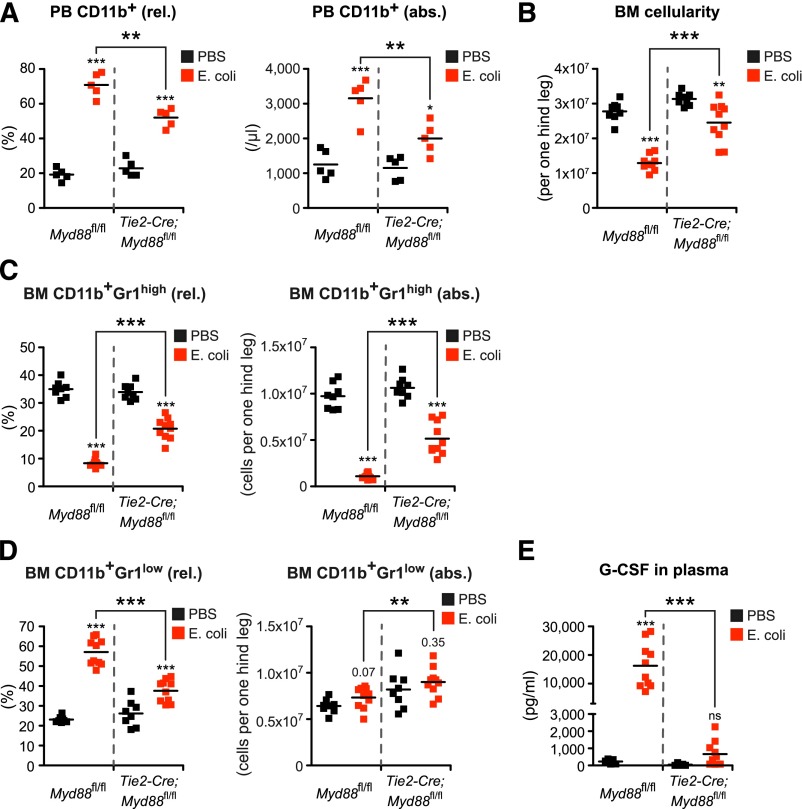
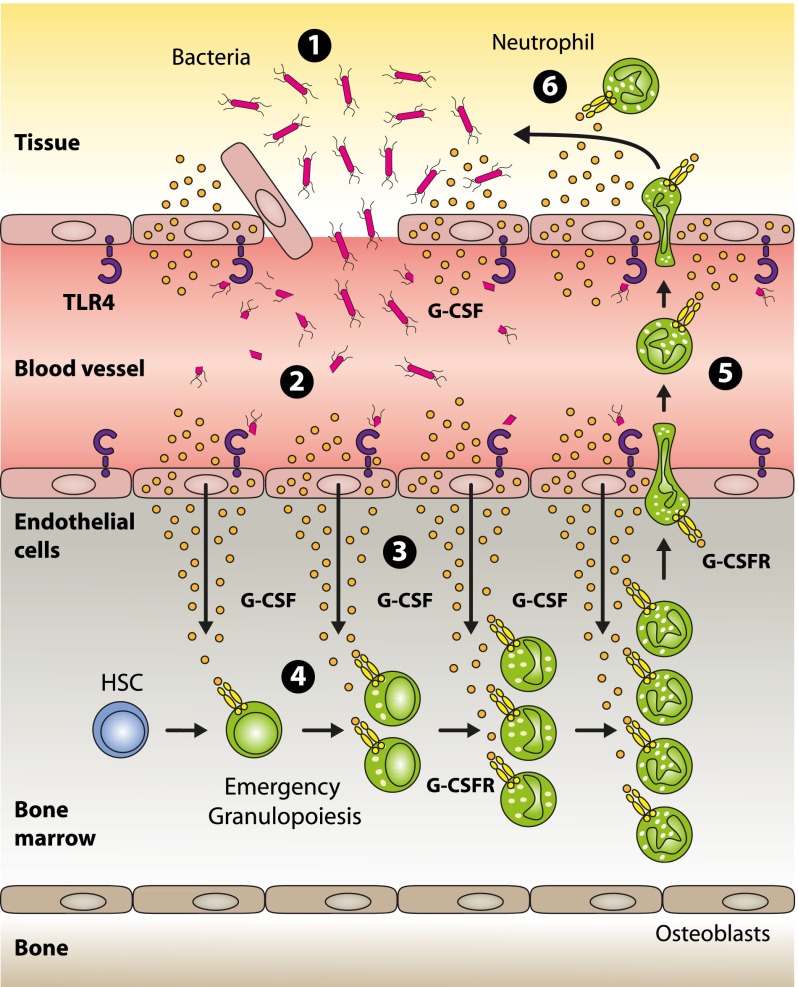
Systemic bacterial infection induces a hematopoietic response program termed "emergency granulopoiesis" that is characterized by increased de novo bone marrow (BM) neutrophil production. How loss of local immune control and bacterial dissemination is sensed and subsequently translated into the switch from steady-state to emergency granulopoiesis is, however, unknown. Using tissue-specific myeloid differentiation primary response gene 88 (Myd88)-deficient mice and in vivo lipopolysaccharide (LPS) administration to model severe bacterial infection, we here show that endothelial cells (ECs) but not hematopoietic cells, hepatocytes, pericytes, or BM stromal cells, are essential cells for this process. Indeed, ECs from multiple tissues including BM express high levels of Tlr4 and Myd88 and are the primary source of granulocyte colony-stimulating factor (G-CSF), the key granulopoietic cytokine, after LPS challenge or infection with Escherichia coli. EC-intrinsic MYD88 signaling and subsequent G-CSF production by ECs is required for myeloid progenitor lineage skewing toward granulocyte-macrophage progenitors, increased colony-forming unit granulocyte activity in BM, and accelerated BM neutrophil generation after LPS stimulation. Thus, ECs catalyze the detection of systemic infection into demand-adapted granulopoiesis.
© 2014 by The American Society of Hematology.
Figures
Comment in
-
Microbial messaging to the marrow.Blood. 2014 Aug 28;124(9):1379-80. doi: 10.1182/blood-2014-07-587345. Blood. 2014. PMID: 25170108 Free PMC article.
References
-
- Lieschke GJ, Grail D, Hodgson G, et al. Mice lacking granulocyte colony-stimulating factor have chronic neutropenia, granulocyte and macrophage progenitor cell deficiency, and impaired neutrophil mobilization. Blood. 1994;84(6):1737–1746. - PubMed
-
- Kolaczkowska E, Kubes P. Neutrophil recruitment and function in health and inflammation. Nat Rev Immunol. 2013;13(3):159–175. - PubMed
-
- Elks PM, van Eeden FJ, Dixon G, et al. Activation of hypoxia-inducible factor-1α (Hif-1α) delays inflammation resolution by reducing neutrophil apoptosis and reverse migration in a zebrafish inflammation model. Blood. 2011;118(3):712–722. - PubMed
-
- Mathias JR, Perrin BJ, Liu T-X, Kanki J, Look AT, Huttenlocher A. Resolution of inflammation by retrograde chemotaxis of neutrophils in transgenic zebrafish. J Leukoc Biol. 2006;80(6):1281–1288. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
