

Heterochromatin-associated interactions of Drosophila HP1a with dADD1, HIPP1, and repetitive RNAs
- PMID: 24990964
- PMCID: PMC4083088
- DOI: 10.1101/gad.241950.114
Heterochromatin-associated interactions of Drosophila HP1a with dADD1, HIPP1, and repetitive RNAs
Abstract
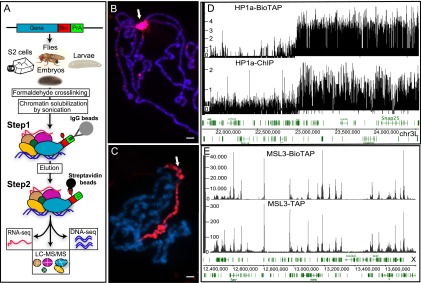
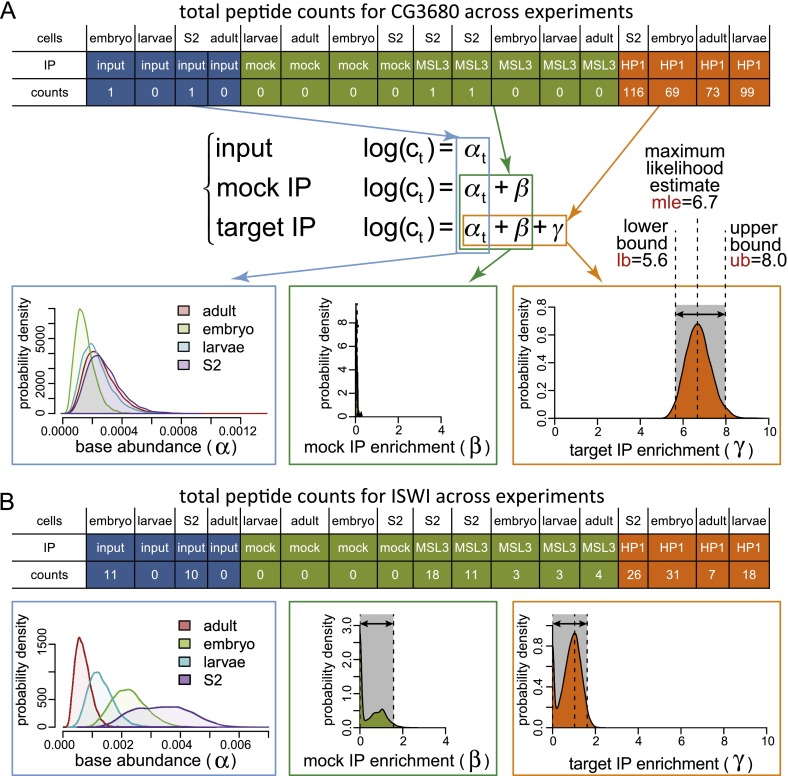
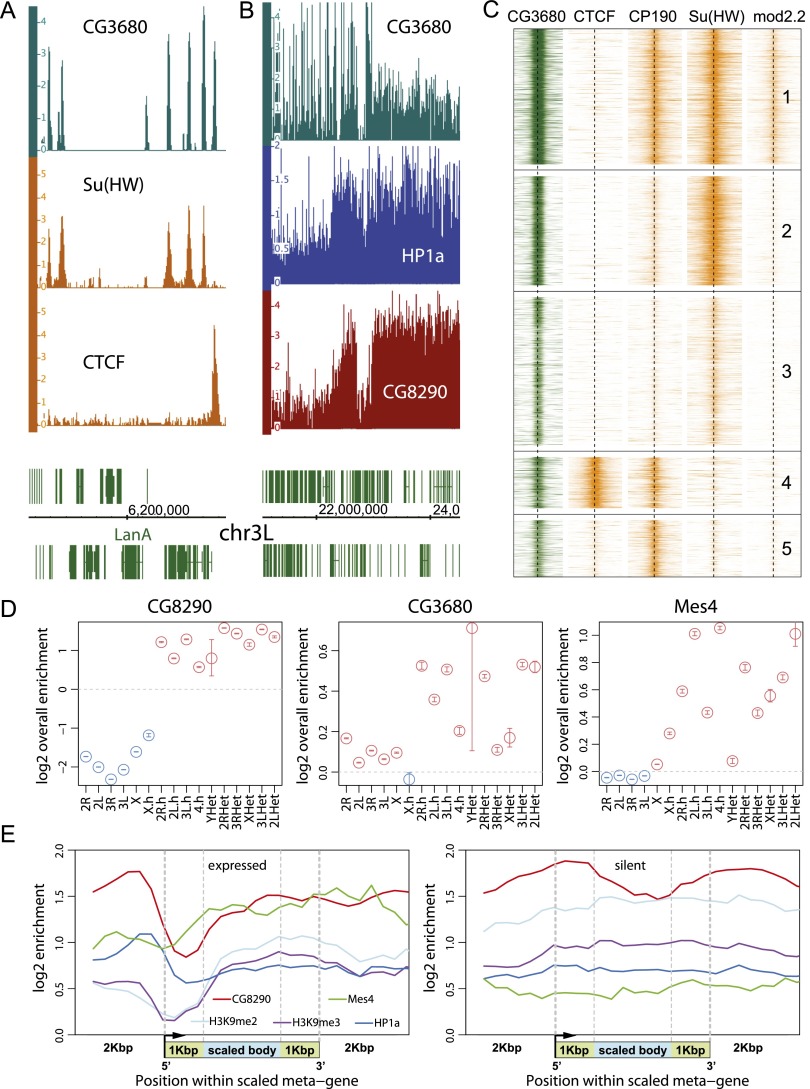
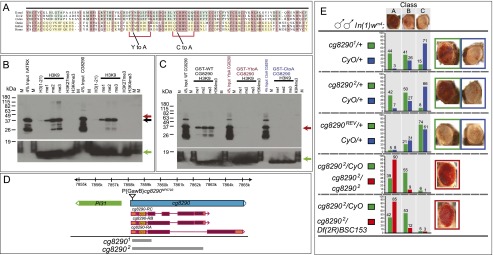
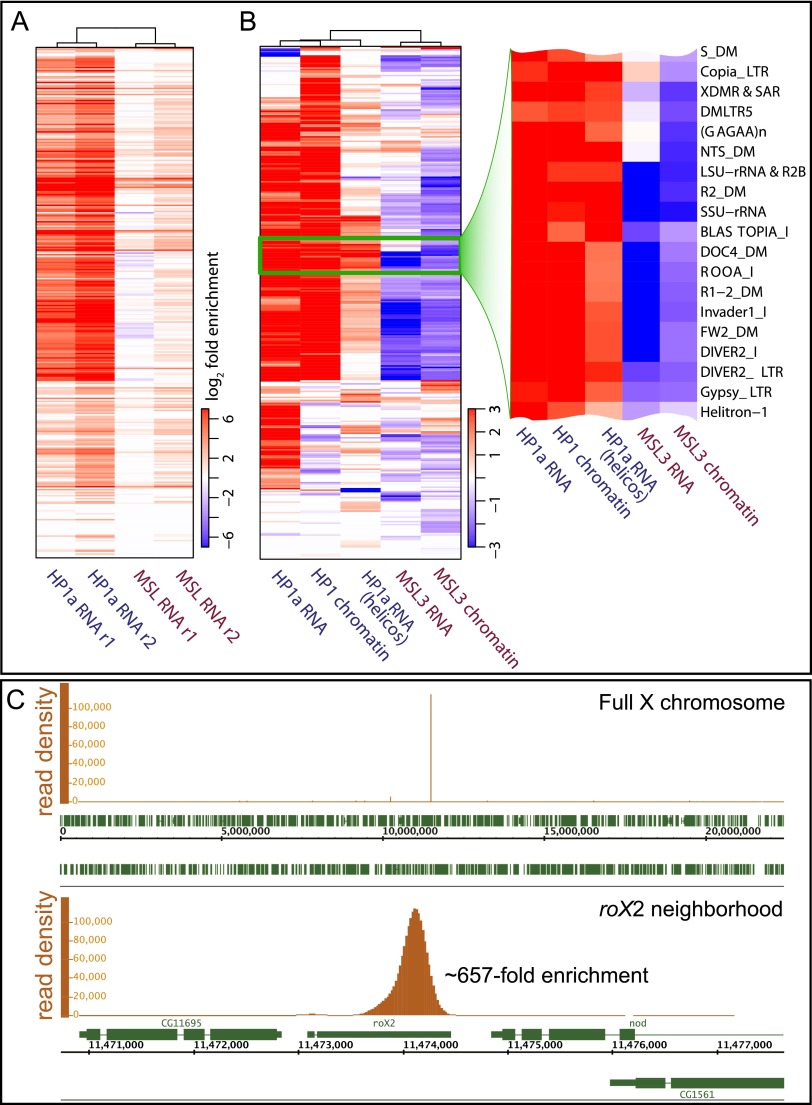
Heterochromatin protein 1 (HP1a) has conserved roles in gene silencing and heterochromatin and is also implicated in transcription, DNA replication, and repair. Here we identify chromatin-associated protein and RNA interactions of HP1a by BioTAP-XL mass spectrometry and sequencing from Drosophila S2 cells, embryos, larvae, and adults. Our results reveal an extensive list of known and novel HP1a-interacting proteins, of which we selected three for validation. A strong novel interactor, dADD1 (Drosophila ADD1) (CG8290), is highly enriched in heterochromatin, harbors an ADD domain similar to human ATRX, displays selective binding to H3K9me2 and H3K9me3, and is a classic genetic suppressor of position-effect variegation. Unexpectedly, a second hit, HIPP1 (HP1 and insulator partner protein-1) (CG3680), is strongly connected to CP190-related complexes localized at putative insulator sequences throughout the genome in addition to its colocalization with HP1a in heterochromatin. A third interactor, the histone methyltransferase MES-4, is also enriched in heterochromatin. In addition to these protein-protein interactions, we found that HP1a selectively associated with a broad set of RNAs transcribed from repetitive regions. We propose that this rich network of previously undiscovered interactions will define how HP1a complexes perform their diverse functions in cells and developing organisms.
Keywords: Bayesian analysis; ChIP; HP1a; LC-MS/MS; formaldehyde cross-linking.
© 2014 Alekseyenko et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Amrein H, Axel R 1997. Genes expressed in neurons of adult male Drosophila. Cell 88: 459–469 - PubMed
-
- Ashburner M (1989). Drosophila, A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous