

Ontogeny of object versus location recognition in the rat: acquisition and retention effects
- PMID: 24992011
- PMCID: PMC4786941
- DOI: 10.1002/dev.21232
Ontogeny of object versus location recognition in the rat: acquisition and retention effects
Abstract
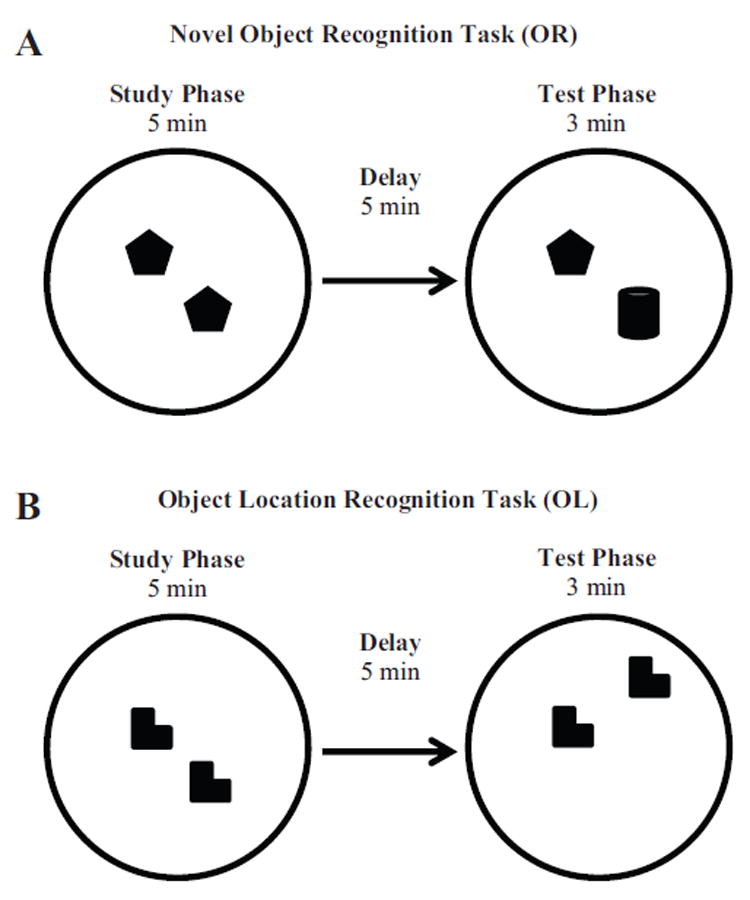
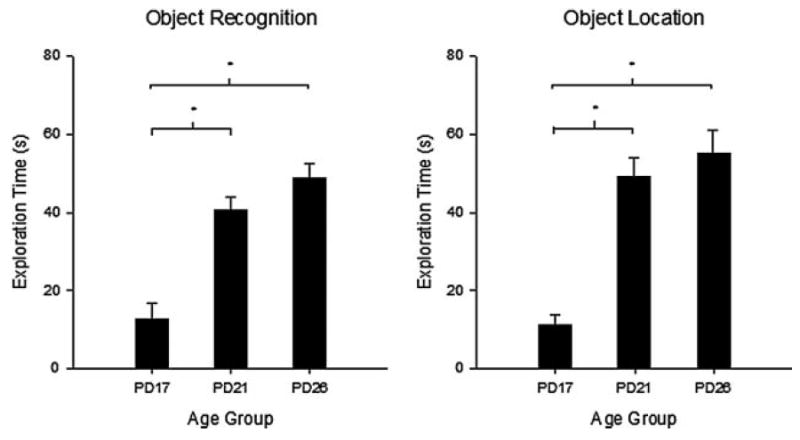
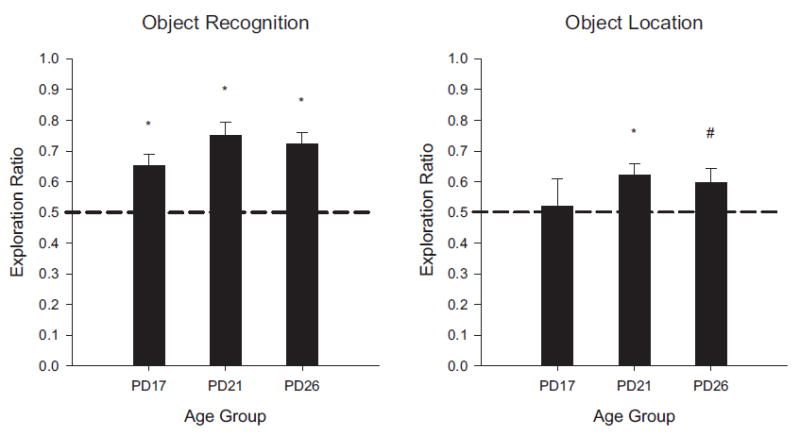
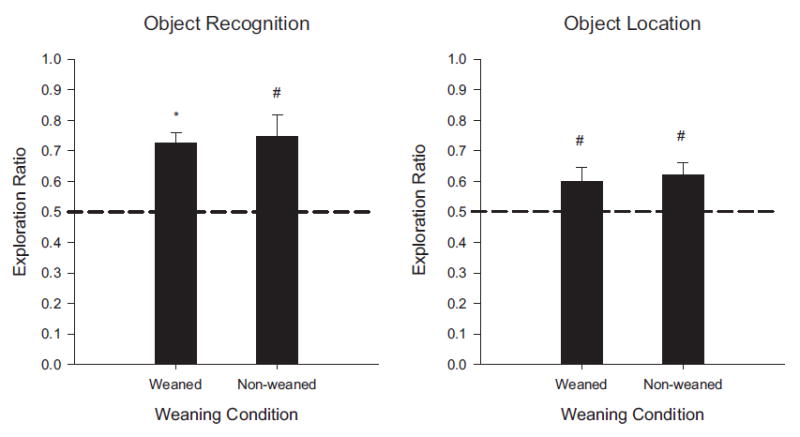
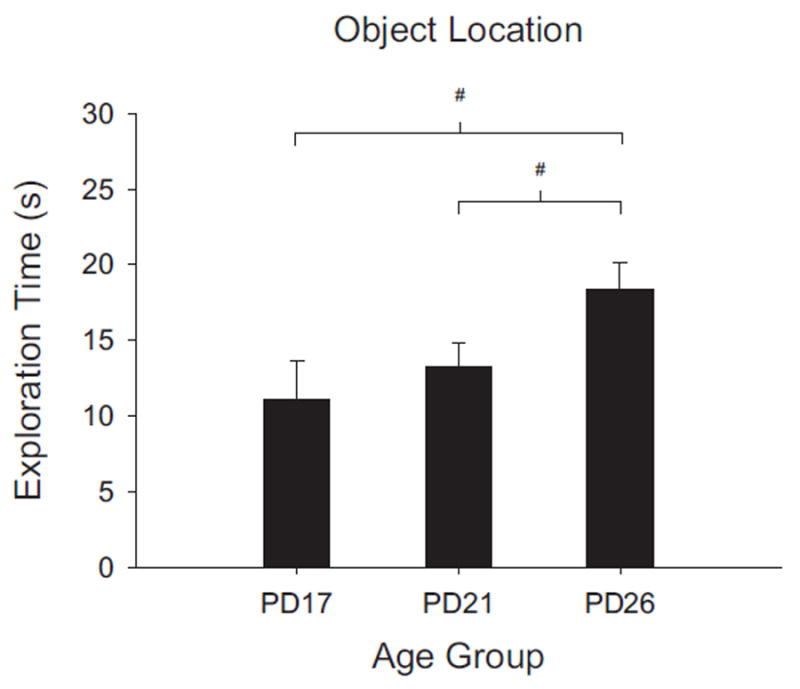
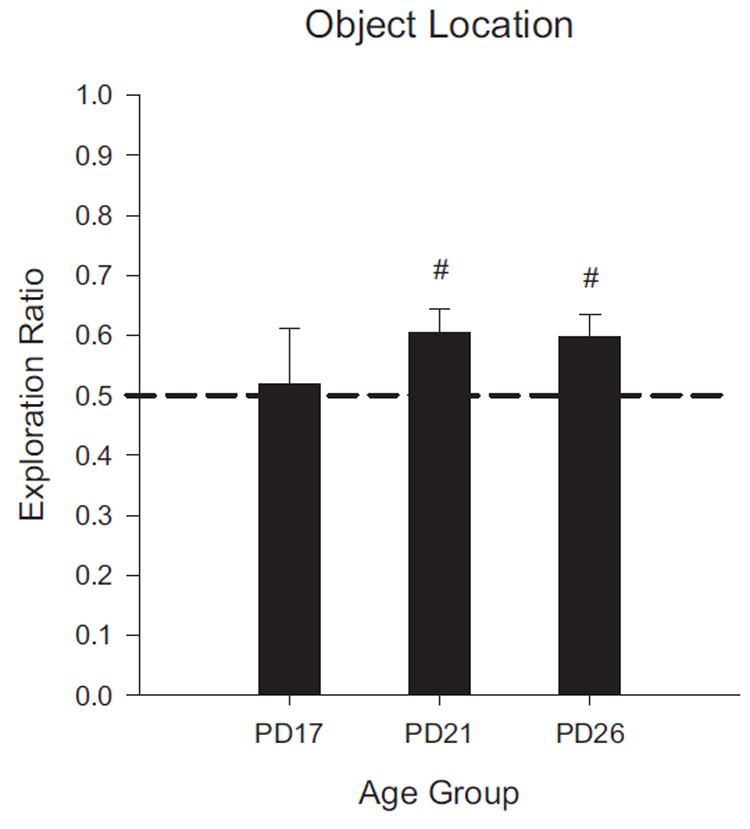
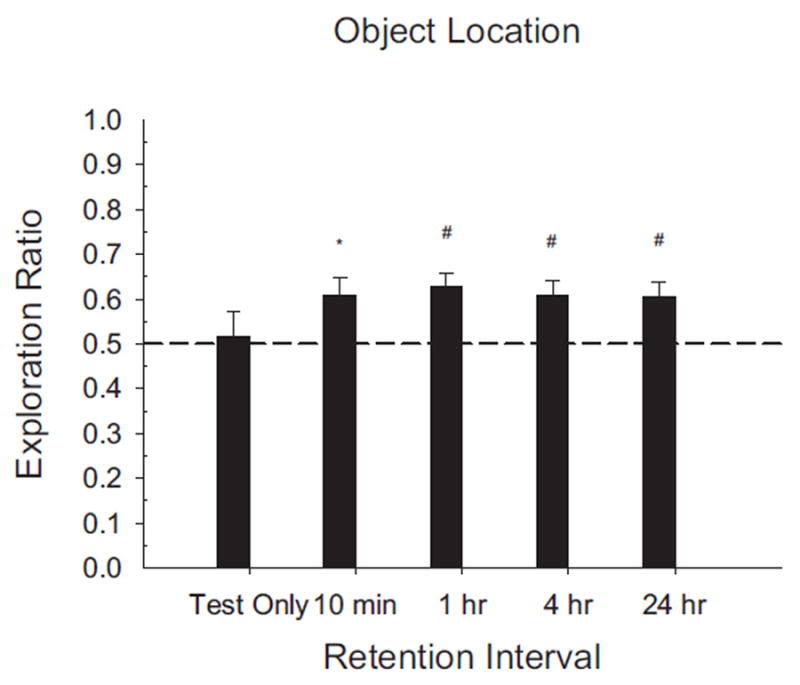
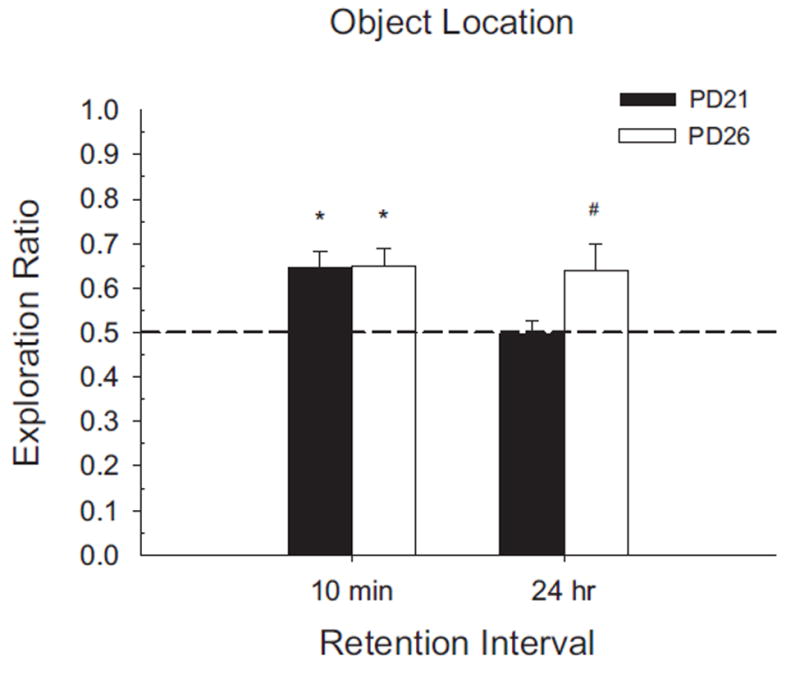
Novel object and location recognition tasks harness the rat's natural tendency to explore novelty (Berlyne, 1950) to study incidental learning. The present study examined the ontogenetic profile of these two tasks and retention of spatial learning between postnatal day (PD) 17 and 31. Experiment 1 showed that rats ages PD17, 21, and 26 recognize novel objects, but only PD21 and PD26 rats recognize a novel location of a familiar object. These results suggest that novel object recognition develops before PD17, while object location recognition emerges between PD17 and PD21. Experiment 2 studied the ontogenetic profile of object location memory retention in PD21, 26, and 31 rats. PD26 and PD31 rats retained the object location memory for both 10-min and 24-hr delays. PD21 rats failed to retain the object location memory for the 24-hr delay, suggesting differential development of short- versus long-term memory in the ontogeny of object location memory.
Keywords: Long-Evans; hippocampus; incidental learning; long-term retention; perirhinal cortex; short-term retention.
© 2014 Wiley Periodicals, Inc.
Figures
Similar articles
-
Infant rats can acquire, but not retain contextual associations in object-in-context and contextual fear conditioning paradigms.Dev Psychobiol. 2020 Dec;62(8):1158-1164. doi: 10.1002/dev.21980. Epub 2020 May 22. Dev Psychobiol. 2020. PMID: 32441808
-
Determinants of novel object and location recognition during development.Behav Brain Res. 2013 Nov 1;256:140-50. doi: 10.1016/j.bbr.2013.07.055. Epub 2013 Aug 8. Behav Brain Res. 2013. PMID: 23933466 Free PMC article.
-
Recent and remote retrograde memory deficit in rats with medial entorhinal cortex lesions.Neurobiol Learn Mem. 2018 Nov;155:157-163. doi: 10.1016/j.nlm.2018.07.013. Epub 2018 Jul 31. Neurobiol Learn Mem. 2018. PMID: 30075194 Free PMC article.
-
Ontogeny of spontaneous recognition memory in rodents.Neurobiol Learn Mem. 2021 Jan;177:107361. doi: 10.1016/j.nlm.2020.107361. Epub 2020 Dec 8. Neurobiol Learn Mem. 2021. PMID: 33307181 Review.
-
Regulation of object recognition and object placement by ovarian sex steroid hormones.Behav Brain Res. 2015 May 15;285:140-57. doi: 10.1016/j.bbr.2014.08.001. Epub 2014 Aug 15. Behav Brain Res. 2015. PMID: 25131507 Free PMC article. Review.
Cited by
-
Developmental trajectory of episodic-like memory in rats.Front Behav Neurosci. 2022 Nov 29;16:969871. doi: 10.3389/fnbeh.2022.969871. eCollection 2022. Front Behav Neurosci. 2022. PMID: 36523755 Free PMC article.
-
Spontaneous Object Exploration in a Recessive Gene Knockout Model of Parkinson's Disease: Development and Progression of Object Recognition Memory Deficits in Male Pink1-/- Rats.Front Behav Neurosci. 2022 Dec 6;16:951268. doi: 10.3389/fnbeh.2022.951268. eCollection 2022. Front Behav Neurosci. 2022. PMID: 36560930 Free PMC article.
-
Development of working memory in the male adolescent rat.Dev Cogn Neurosci. 2019 Jun;37:100601. doi: 10.1016/j.dcn.2018.11.003. Epub 2018 Nov 22. Dev Cogn Neurosci. 2019. PMID: 30497917 Free PMC article.
-
Cortico-Hippocampal Oscillations Are Associated With the Developmental Onset of Hippocampal-Dependent Memory.Front Neurosci. 2022 Jun 23;16:891523. doi: 10.3389/fnins.2022.891523. eCollection 2022. Front Neurosci. 2022. PMID: 35812209 Free PMC article.
-
Extended access self-administration of methamphetamine is associated with age- and sex-dependent differences in drug taking behavior and recognition memory in rats.Behav Brain Res. 2020 Jul 15;390:112659. doi: 10.1016/j.bbr.2020.112659. Epub 2020 May 8. Behav Brain Res. 2020. PMID: 32437887 Free PMC article.
References
-
- Bayer SA, Altman J. Hippocampal development in the rat: Cytogenesis and morphogenesis examined with autoradiography and low-level X-irradiation. Journal of Comparative Neurology. 1978;158(1):55–79. - PubMed
-
- Berlyne DE. Novelty and curiosity as determinants of exploratory behavior. British Journal of Psychology. 1950;41:68.
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
