

Genetic analysis of Leishmania donovani tropism using a naturally attenuated cutaneous strain
- PMID: 24992200
- PMCID: PMC4081786
- DOI: 10.1371/journal.ppat.1004244
Genetic analysis of Leishmania donovani tropism using a naturally attenuated cutaneous strain
Abstract
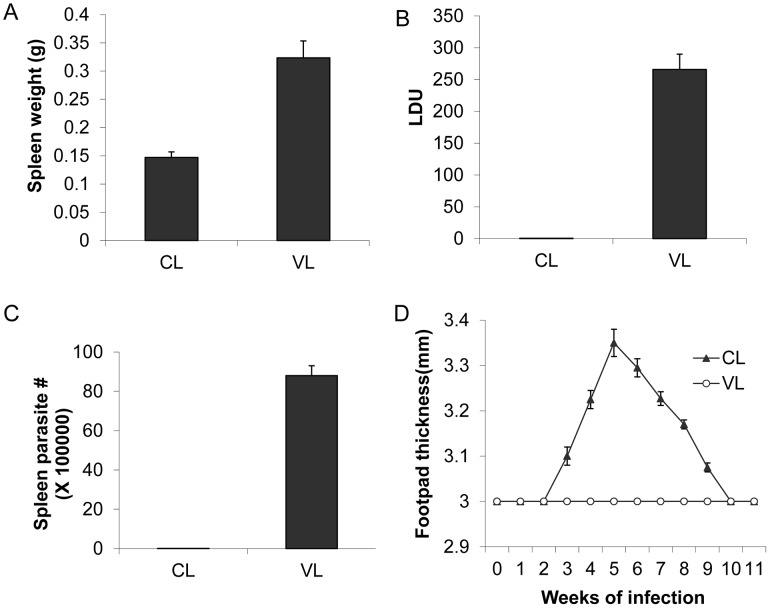
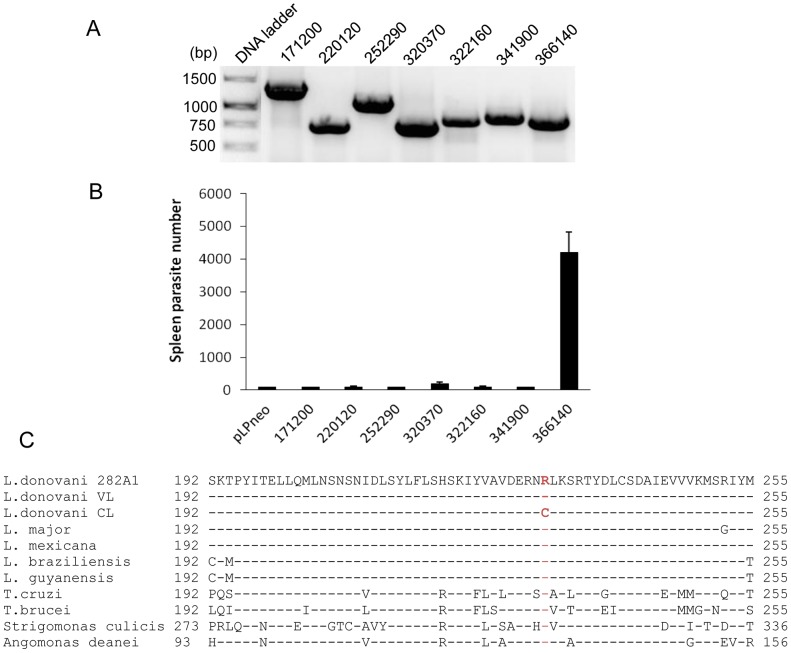
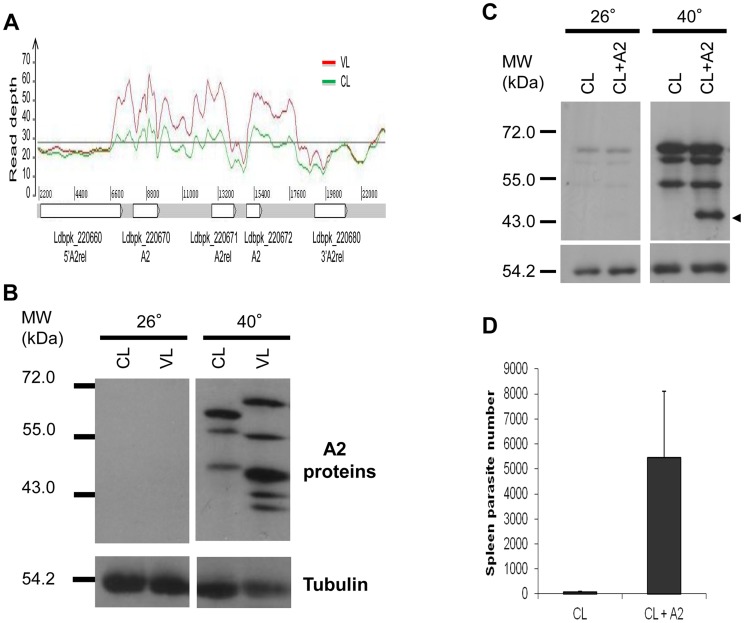
A central question in Leishmania research is why most species cause cutaneous infections but others cause fatal visceral disease. Interestingly, L. donovani causes both visceral and cutaneous leishmaniasis in Sri Lanka. L. donovani clinical isolates were therefore obtained from cutaneous leishmaniasis (CL-SL) and visceral leishmaniasis (VL-SL) patients from Sri Lanka. The CL-SL isolate was severely attenuated compared to the VL-SL isolate for survival in visceral organs in BALB/c mice. Genomic and transcriptomic analysis argue that gene deletions or pseudogenes specific to CL-SL are not responsible for the difference in disease tropism and that single nucleotide polymorphisms (SNPs) and/or gene copy number variations play a major role in altered pathology. This is illustrated through the observations within showing that a decreased copy number of the A2 gene family and a mutation in the ras-like RagC GTPase enzyme in the mTOR pathway contribute to the attenuation of the CL-SL strain in visceral infection. Overall, this research provides a unique perspective on genetic differences associated with diverse pathologies caused by Leishmania infection.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Similar articles
-
Evidence that a naturally occurring single nucleotide polymorphism in the RagC gene of Leishmania donovani contributes to reduced virulence.PLoS Negl Trop Dis. 2021 Feb 23;15(2):e0009079. doi: 10.1371/journal.pntd.0009079. eCollection 2021 Feb. PLoS Negl Trop Dis. 2021. PMID: 33621241 Free PMC article.
-
Genomic insights into virulence mechanisms of Leishmania donovani: evidence from an atypical strain.BMC Genomics. 2018 Nov 28;19(1):843. doi: 10.1186/s12864-018-5271-z. BMC Genomics. 2018. PMID: 30486770 Free PMC article.
-
Comparison of the A2 gene locus in Leishmania donovani and Leishmania major and its control over cutaneous infection.J Biol Chem. 2003 Sep 12;278(37):35508-15. doi: 10.1074/jbc.M305030200. Epub 2003 Jun 26. J Biol Chem. 2003. PMID: 12829719
-
PKDL and other dermal lesions in HIV co-infected patients with Leishmaniasis: review of clinical presentation in relation to immune responses.PLoS Negl Trop Dis. 2014 Nov 20;8(11):e3258. doi: 10.1371/journal.pntd.0003258. eCollection 2014. PLoS Negl Trop Dis. 2014. PMID: 25412435 Free PMC article. Review.
-
Determinants for the development of visceral leishmaniasis disease.PLoS Pathog. 2013 Jan;9(1):e1003053. doi: 10.1371/journal.ppat.1003053. Epub 2013 Jan 3. PLoS Pathog. 2013. PMID: 23300451 Free PMC article. Review.
Cited by
-
A complete Leishmania donovani reference genome identifies novel genetic variations associated with virulence.Sci Rep. 2018 Nov 8;8(1):16549. doi: 10.1038/s41598-018-34812-x. Sci Rep. 2018. PMID: 30409989 Free PMC article.
-
Identification of divergent Leishmania (Viannia) braziliensis ecotypes derived from a geographically restricted area through whole genome analysis.PLoS Negl Trop Dis. 2019 Jun 6;13(6):e0007382. doi: 10.1371/journal.pntd.0007382. eCollection 2019 Jun. PLoS Negl Trop Dis. 2019. PMID: 31170148 Free PMC article.
-
Leishmania donovani persistence and circulation causing cutaneous leishmaniasis in unusual-foci of Nepal.Sci Rep. 2023 Jul 29;13(1):12329. doi: 10.1038/s41598-023-37458-6. Sci Rep. 2023. PMID: 37516780 Free PMC article.
-
Efficacy of a new rapid diagnostic test kit to diagnose Sri Lankan cutaneous leishmaniasis caused by Leishmania donovani.PLoS One. 2017 Nov 14;12(11):e0187024. doi: 10.1371/journal.pone.0187024. eCollection 2017. PLoS One. 2017. PMID: 29135995 Free PMC article.
-
L-arginine availability and arginase activity: Characterization of amino acid permease 3 in Leishmania amazonensis.PLoS Negl Trop Dis. 2017 Oct 26;11(10):e0006025. doi: 10.1371/journal.pntd.0006025. eCollection 2017 Oct. PLoS Negl Trop Dis. 2017. PMID: 29073150 Free PMC article.
References
-
- Murray HW, Berman JD, Davies CR, Saravia NG (2005) Advances in leishmaniasis. Lancet 366: 1561–1577. - PubMed
-
- Karunaweera ND, Pratlong F, Siriwardane HVYD, Ihalamulla RL, Dedet JP (2003) Lankan cutaneous leishmaniasis is caused by Leishmania donovani zymodeme MON-37. Transactions of the Royal Society of Tropical Medicine and Hygiene 97: 380–381. - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
