

Meiosis-specific cohesin component, Stag3 is essential for maintaining centromere chromatid cohesion, and required for DNA repair and synapsis between homologous chromosomes
- PMID: 24992337
- PMCID: PMC4081007
- DOI: 10.1371/journal.pgen.1004413
Meiosis-specific cohesin component, Stag3 is essential for maintaining centromere chromatid cohesion, and required for DNA repair and synapsis between homologous chromosomes
Abstract
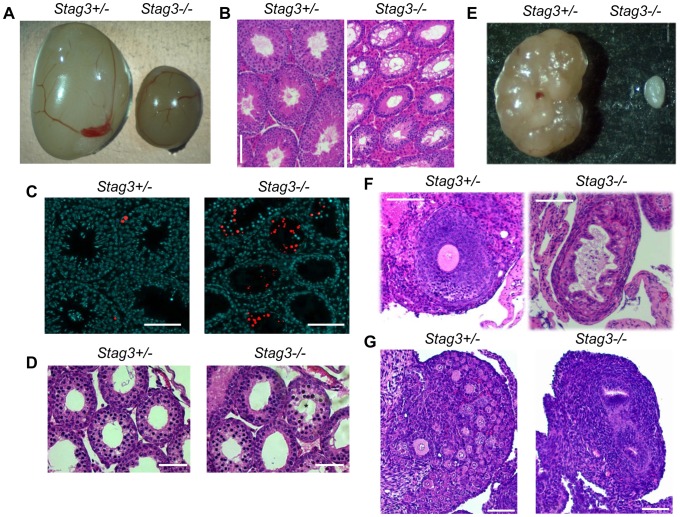
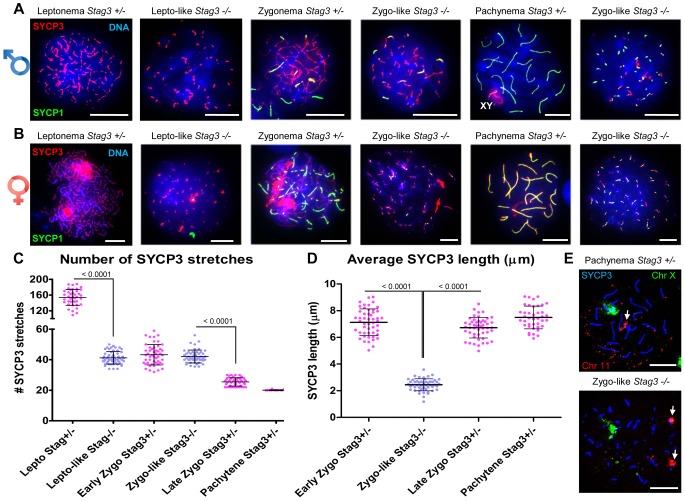
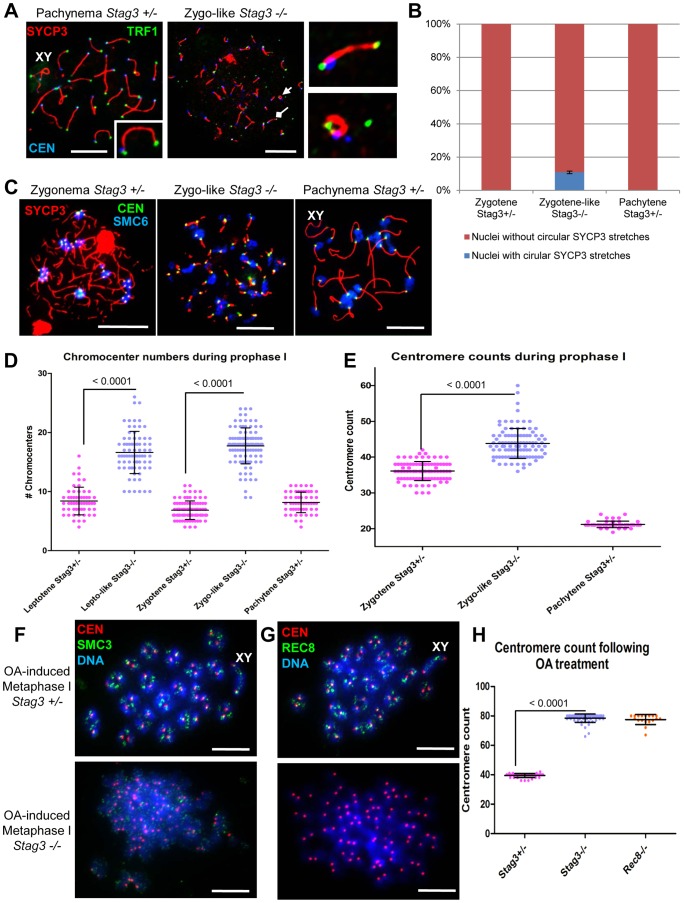
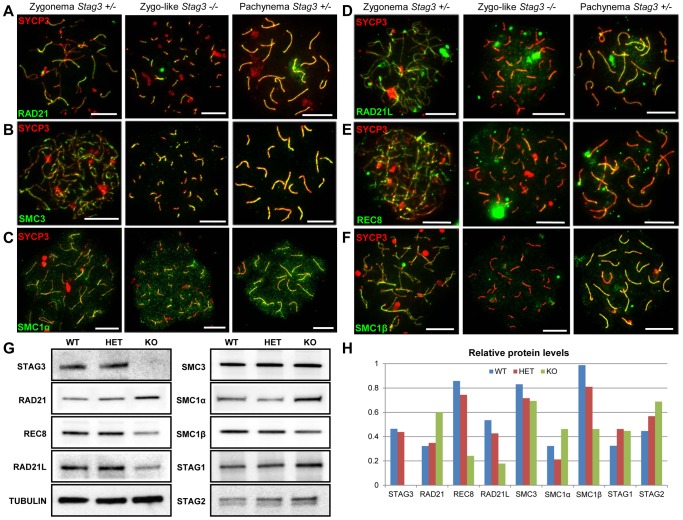
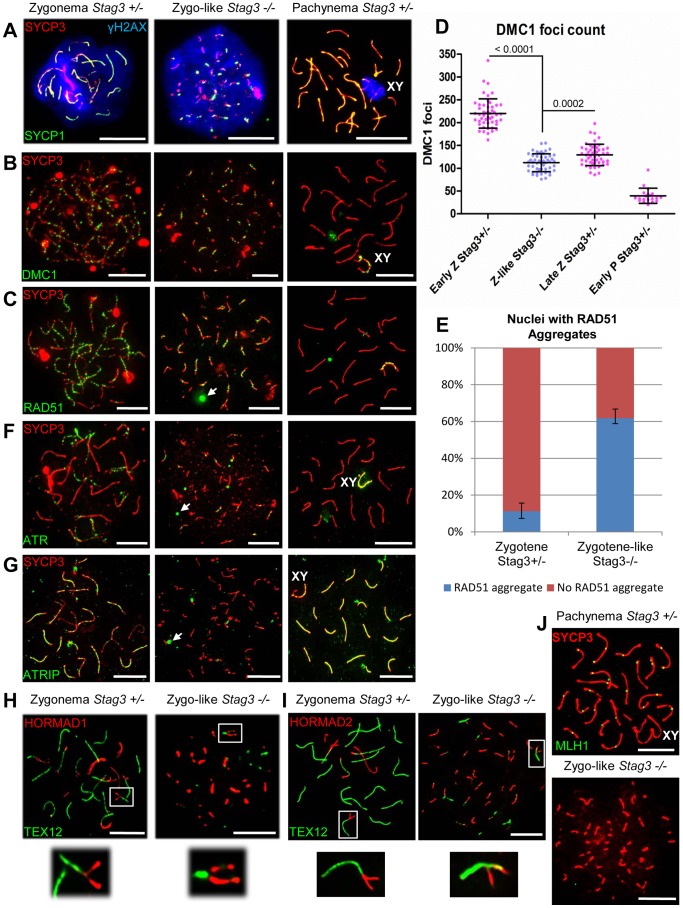
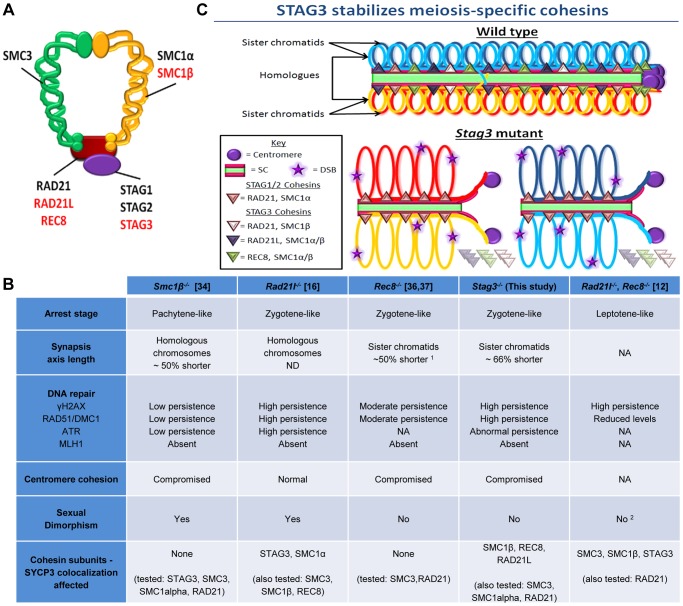
Cohesins are important for chromosome structure and chromosome segregation during mitosis and meiosis. Cohesins are composed of two structural maintenance of chromosomes (SMC1-SMC3) proteins that form a V-shaped heterodimer structure, which is bridged by a α-kleisin protein and a stromal antigen (STAG) protein. Previous studies in mouse have shown that there is one SMC1 protein (SMC1β), two α-kleisins (RAD21L and REC8) and one STAG protein (STAG3) that are meiosis-specific. During meiosis, homologous chromosomes must recombine with one another in the context of a tripartite structure known as the synaptonemal complex (SC). From interaction studies, it has been shown that there are at least four meiosis-specific forms of cohesin, which together with the mitotic cohesin complex, are lateral components of the SC. STAG3 is the only meiosis-specific subunit that is represented within all four meiosis-specific cohesin complexes. In Stag3 mutant germ cells, the protein level of other meiosis-specific cohesin subunits (SMC1β, RAD21L and REC8) is reduced, and their localization to chromosome axes is disrupted. In contrast, the mitotic cohesin complex remains intact and localizes robustly to the meiotic chromosome axes. The instability of meiosis-specific cohesins observed in Stag3 mutants results in aberrant DNA repair processes, and disruption of synapsis between homologous chromosomes. Furthermore, mutation of Stag3 results in perturbation of pericentromeric heterochromatin clustering, and disruption of centromere cohesion between sister chromatids during meiotic prophase. These defects result in early prophase I arrest and apoptosis in both male and female germ cells. The meiotic defects observed in Stag3 mutants are more severe when compared to single mutants for Smc1β, Rec8 and Rad21l, however they are not as severe as the Rec8, Rad21l double mutants. Taken together, our study demonstrates that STAG3 is required for the stability of all meiosis-specific cohesin complexes. Furthermore, our data suggests that STAG3 is required for structural changes of chromosomes that mediate chromosome pairing and synapsis, DNA repair and progression of meiosis.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Genetic Interactions Between the Meiosis-Specific Cohesin Components, STAG3, REC8, and RAD21L.G3 (Bethesda). 2016 Jun 1;6(6):1713-24. doi: 10.1534/g3.116.029462. G3 (Bethesda). 2016. PMID: 27172213 Free PMC article.
-
STAG3-mediated stabilization of REC8 cohesin complexes promotes chromosome synapsis during meiosis.EMBO J. 2014 Jun 2;33(11):1243-55. doi: 10.1002/embj.201387329. Epub 2014 May 5. EMBO J. 2014. PMID: 24797475 Free PMC article.
-
Distinct Roles of Meiosis-Specific Cohesin Complexes in Mammalian Spermatogenesis.PLoS Genet. 2016 Oct 28;12(10):e1006389. doi: 10.1371/journal.pgen.1006389. eCollection 2016 Oct. PLoS Genet. 2016. PMID: 27792785 Free PMC article.
-
Rec8 Cohesin: A Structural Platform for Shaping the Meiotic Chromosomes.Genes (Basel). 2022 Jan 22;13(2):200. doi: 10.3390/genes13020200. Genes (Basel). 2022. PMID: 35205245 Free PMC article. Review.
-
Meiotic prophase-like pathway for cleavage-independent removal of cohesin for chromosome morphogenesis.Curr Genet. 2019 Aug;65(4):817-827. doi: 10.1007/s00294-019-00959-x. Epub 2019 Mar 28. Curr Genet. 2019. PMID: 30923890 Review.
Cited by
-
Stag3 regulates microtubule stability to maintain euploidy during mouse oocyte meiotic maturation.Oncotarget. 2017 Jan 3;8(1):1593-1602. doi: 10.18632/oncotarget.13684. Oncotarget. 2017. PMID: 27906670 Free PMC article.
-
Complex elaboration: making sense of meiotic cohesin dynamics.FEBS J. 2015 Jul;282(13):2426-43. doi: 10.1111/febs.13301. Epub 2015 May 9. FEBS J. 2015. PMID: 25895170 Free PMC article. Review.
-
SMC1α Substitutes for Many Meiotic Functions of SMC1β but Cannot Protect Telomeres from Damage.Curr Biol. 2018 Jan 22;28(2):249-261.e4. doi: 10.1016/j.cub.2017.12.020. Epub 2018 Jan 11. Curr Biol. 2018. PMID: 29337080 Free PMC article.
-
Regulation of Meiotic Prophase One in Mammalian Oocytes.Front Cell Dev Biol. 2021 May 20;9:667306. doi: 10.3389/fcell.2021.667306. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34095134 Free PMC article. Review.
-
Structural and functional adaptations of the mammalian nuclear envelope to meet the meiotic requirements.Nucleus. 2015;6(2):93-101. doi: 10.1080/19491034.2015.1004941. Epub 2015 Feb 12. Nucleus. 2015. PMID: 25674669 Free PMC article. Review.
References
-
- Jordan P (2006) Initiation of homologous chromosome pairing during meiosis. Biochem Soc Trans 34: 545–549. - PubMed
-
- Nasmyth K (2011) Cohesin: a catenase with separate entry and exit gates? Nat Cell Biol 13: 1170–1177. - PubMed
-
- Prieto I, Suja JA, Pezzi N, Kremer L, Martinez-A C, et al. (2001) Mammalian STAG3 is a cohesin specific to sister chromatid arms in meiosis I. Nat Cell Biol 3: 761–766. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
