

Regulation of the nucleosome repeat length in vivo by the DNA sequence, protein concentrations and long-range interactions
- PMID: 24992723
- PMCID: PMC4081033
- DOI: 10.1371/journal.pcbi.1003698
Regulation of the nucleosome repeat length in vivo by the DNA sequence, protein concentrations and long-range interactions
Abstract
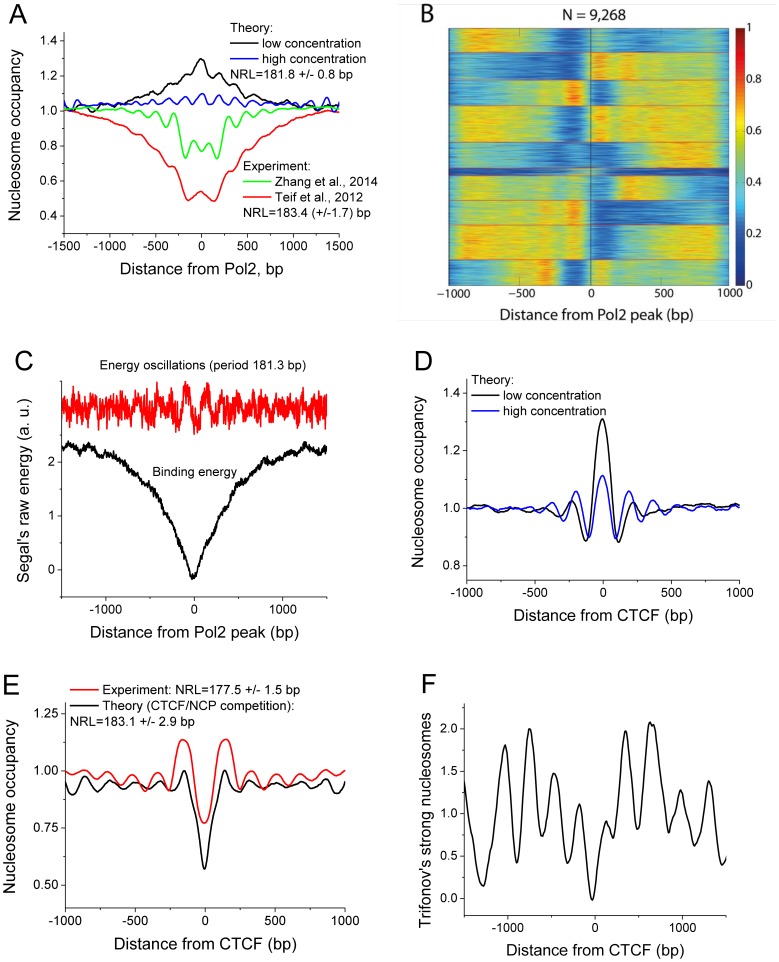
The nucleosome repeat length (NRL) is an integral chromatin property important for its biological functions. Recent experiments revealed several conflicting trends of the NRL dependence on the concentrations of histones and other architectural chromatin proteins, both in vitro and in vivo, but a systematic theoretical description of NRL as a function of DNA sequence and epigenetic determinants is currently lacking. To address this problem, we have performed an integrative biophysical and bioinformatics analysis in species ranging from yeast to frog to mouse where NRL was studied as a function of various parameters. We show that in simple eukaryotes such as yeast, a lower limit for the NRL value exists, determined by internucleosome interactions and remodeler action. For higher eukaryotes, also the upper limit exists since NRL is an increasing but saturating function of the linker histone concentration. Counterintuitively, smaller H1 variants or non-histone architectural proteins can initiate larger effects on the NRL due to entropic reasons. Furthermore, we demonstrate that different regimes of the NRL dependence on histone concentrations exist depending on whether DNA sequence-specific effects dominate over boundary effects or vice versa. We consider several classes of genomic regions with apparently different regimes of the NRL variation. As one extreme, our analysis reveals that the period of oscillations of the nucleosome density around bound RNA polymerase coincides with the period of oscillations of positioning sites of the corresponding DNA sequence. At another extreme, we show that although mouse major satellite repeats intrinsically encode well-defined nucleosome preferences, they have no unique nucleosome arrangement and can undergo a switch between two distinct types of nucleosome positioning.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


Similar articles
-
A comparison of in vitro nucleosome positioning mapped with chicken, frog and a variety of yeast core histones.J Mol Biol. 2013 Nov 15;425(22):4206-22. doi: 10.1016/j.jmb.2013.07.019. Epub 2013 Jul 18. J Mol Biol. 2013. PMID: 23871836 Free PMC article.
-
Role of linker histone in chromatin structure and function: H1 stoichiometry and nucleosome repeat length.Chromosome Res. 2006;14(1):17-25. doi: 10.1007/s10577-005-1024-3. Chromosome Res. 2006. PMID: 16506093 Review.
-
Chromatin fiber polymorphism triggered by variations of DNA linker lengths.Proc Natl Acad Sci U S A. 2014 Jun 3;111(22):8061-6. doi: 10.1073/pnas.1315872111. Epub 2014 May 20. Proc Natl Acad Sci U S A. 2014. PMID: 24847063 Free PMC article.
-
Short nucleosome repeats impose rotational modulations on chromatin fibre folding.EMBO J. 2012 May 16;31(10):2416-26. doi: 10.1038/emboj.2012.80. Epub 2012 Mar 30. EMBO J. 2012. PMID: 22473209 Free PMC article.
-
Emerging roles of linker histones in regulating chromatin structure and function.Nat Rev Mol Cell Biol. 2018 Mar;19(3):192-206. doi: 10.1038/nrm.2017.94. Epub 2017 Oct 11. Nat Rev Mol Cell Biol. 2018. PMID: 29018282 Free PMC article. Review.
Cited by
-
Nucleosome-induced homology recognition in chromatin.J R Soc Interface. 2021 Jun;18(179):20210147. doi: 10.1098/rsif.2021.0147. Epub 2021 Jun 16. J R Soc Interface. 2021. PMID: 34129789 Free PMC article.
-
Why Is a High Temperature Needed by Thermus thermophilus Argonaute During mRNA Silencing: A Theoretical Study.Front Chem. 2018 Jun 14;6:223. doi: 10.3389/fchem.2018.00223. eCollection 2018. Front Chem. 2018. PMID: 29967763 Free PMC article.
-
Nucleosome repositioning during differentiation of a human myeloid leukemia cell line.Nucleus. 2017 Mar 4;8(2):188-204. doi: 10.1080/19491034.2017.1295201. Nucleus. 2017. PMID: 28406749 Free PMC article.
-
One-Dimensional Search Dynamics of Tumor Suppressor p53 Regulated by a Disordered C-Terminal Domain.Biophys J. 2017 Jun 6;112(11):2301-2314. doi: 10.1016/j.bpj.2017.04.038. Biophys J. 2017. PMID: 28591603 Free PMC article.
-
Nucleosomal signatures impose nucleosome positioning in coding and noncoding sequences in the genome.Genome Res. 2016 Nov;26(11):1532-1543. doi: 10.1101/gr.207241.116. Epub 2016 Sep 23. Genome Res. 2016. PMID: 27662899 Free PMC article.
References
-
- van Holde KE (1989) Chromatin. New York: Springer-Verlag. 497 p.
-
- Olins AL, Olins DE (1974) Spheroid chromatin units (v bodies). Science 183: 330–332. - PubMed
-
- Kornberg RD (1974) Chromatin structure: a repeating unit of histones and DNA. Science 184: 868–871. - PubMed
-
- Lohr D, Tatchell K, Van Holde KE (1977) On the occurrence of nucleosome phasing in chromatin. Cell 12: 829–836. - PubMed
-
- Gottesfeld JM, Melton DA (1978) The length of nucleosome-associated DNA is the same in both transcribed and nontranscribed regions of chromatin. Nature 273: 317–319. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
