

Establishment of epithelial polarity--GEF who's minding the GAP?
- PMID: 24994932
- PMCID: PMC4117226
- DOI: 10.1242/jcs.153197
Establishment of epithelial polarity--GEF who's minding the GAP?
Abstract
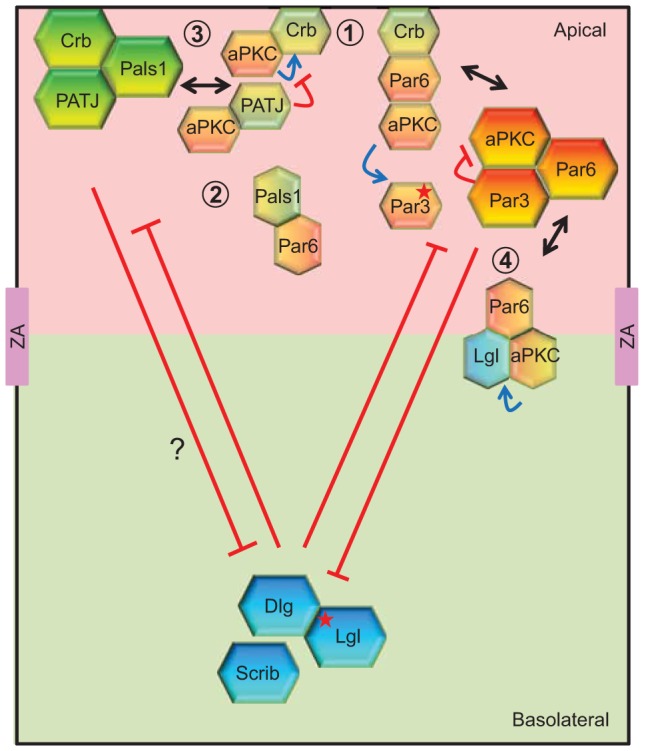
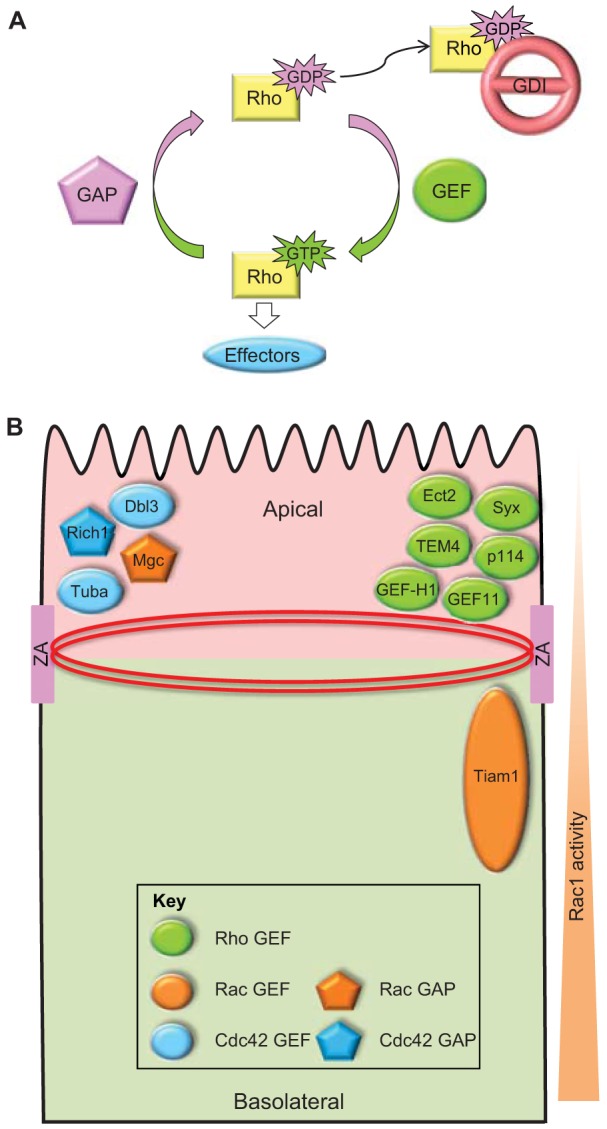
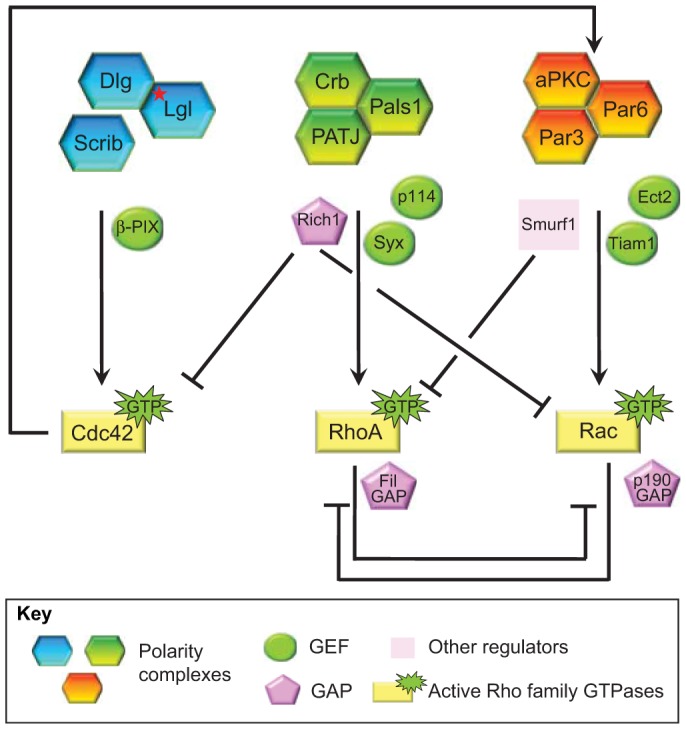
Cell polarization is a fundamental process that underlies epithelial morphogenesis, cell motility, cell division and organogenesis. Loss of polarity predisposes tissues to developmental disorders and contributes to cancer progression. The formation and establishment of epithelial cell polarity is mediated by the cooperation of polarity protein complexes, namely the Crumbs, partitioning defective (Par) and Scribble complexes, with Rho family GTPases, including RhoA, Rac1 and Cdc42. The activation of different GTPases triggers distinct downstream signaling pathways to modulate protein-protein interactions and cytoskeletal remodeling. The spatio-temporal activation and inactivation of these small GTPases is tightly controlled by a complex interconnected network of different regulatory proteins, including guanine-nucleotide-exchange factors (GEFs), GTPase-activating proteins (GAPs), and guanine-nucleotide-dissociation inhibitors (GDIs). In this Commentary, we focus on current understanding on how polarity complexes interact with GEFs and GAPs to control the precise location and activation of Rho GTPases (Crumbs for RhoA, Par for Rac1, and Scribble for Cdc42) to promote apical-basal polarization in mammalian epithelial cells. The mutual exclusion of GTPase activities, especially that of RhoA and Rac1, which is well established, provides a mechanism through which polarity complexes that act through distinct Rho GTPases function as cellular rheostats to fine-tune specific downstream pathways to differentiate and preserve the apical and basolateral domains. This article is part of a Minifocus on Establishing polarity.
Keywords: Cell adhesion; Cell polarity; Crumbs; Par3; Rho GTPase; Scribble.
© 2014. Published by The Company of Biologists Ltd.
Figures
Comment in
-
ERM proteins at a glance.J Cell Sci. 2014 Aug 1;127(Pt 15):3199-204. doi: 10.1242/jcs.098343. Epub 2014 Jun 20. J Cell Sci. 2014. PMID: 24951115 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
