

The importance of microbes in animal development: lessons from the squid-vibrio symbiosis
- PMID: 24995875
- PMCID: PMC6281398
- DOI: 10.1146/annurev-micro-091313-103654
The importance of microbes in animal development: lessons from the squid-vibrio symbiosis
Abstract
Developmental biology is among the many subdisciplines of the life sciences being transformed by our increasing awareness of the role of coevolved microbial symbionts in health and disease. Most symbioses are horizontally acquired, i.e., they begin anew each generation. In such associations, the embryonic period prepares the animal to engage with the coevolved partner(s) with fidelity following birth or hatching. Once interactions are underway, the microbial partners drive maturation of tissues that are either directly associated with or distant from the symbiont populations. Animal alliances often involve complex microbial communities, such as those in the vertebrate gastrointestinal tract. A series of simpler-model systems is providing insight into the basic rules and principles that govern the establishment and maintenance of stable animal-microbe partnerships. This review focuses on what biologists have learned about the developmental trajectory of horizontally acquired symbioses through the study of the binary squid-vibrio model.
Keywords: Euprymna scolopes; Vibrio fischeri; colonization; holobiont; horizontal transmission; morphogenesis.
Conflict of interest statement
DISCLOSURE STATEMENT
The author is not aware of any affiliations, memberships, funding, or financial holdings that might be perceived as affecting the objectivity of this review.
Figures
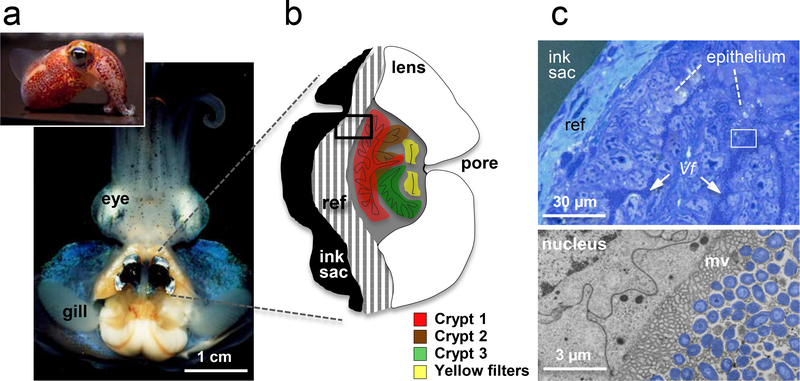
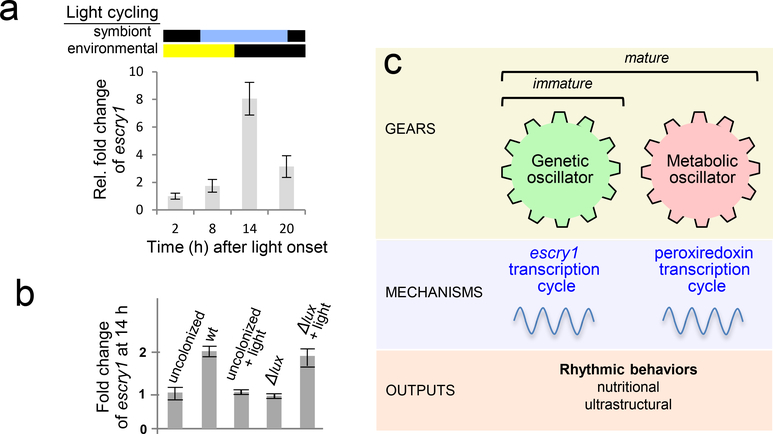
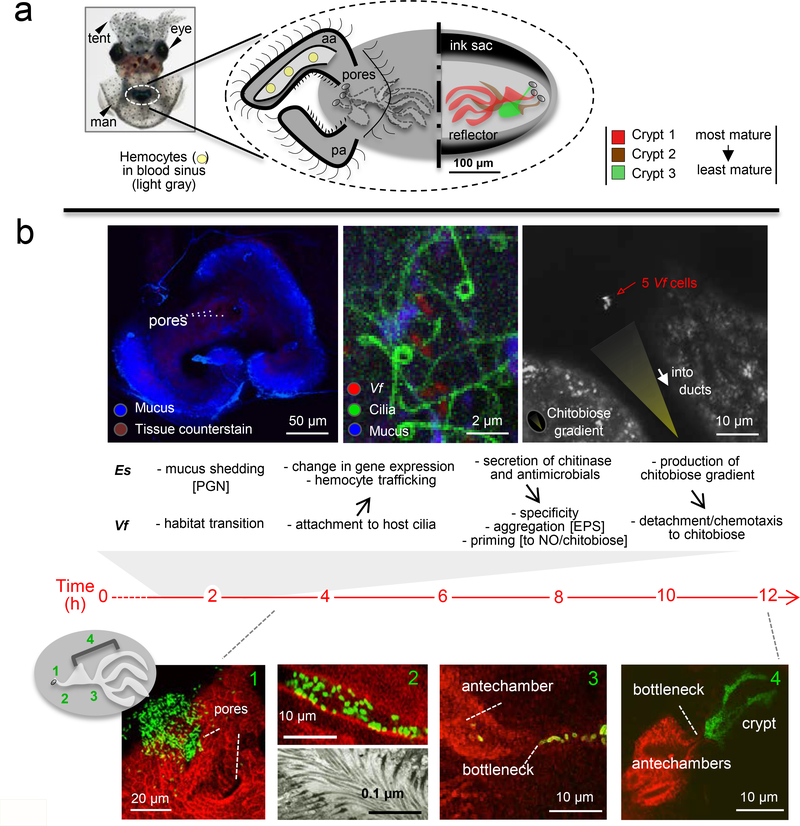
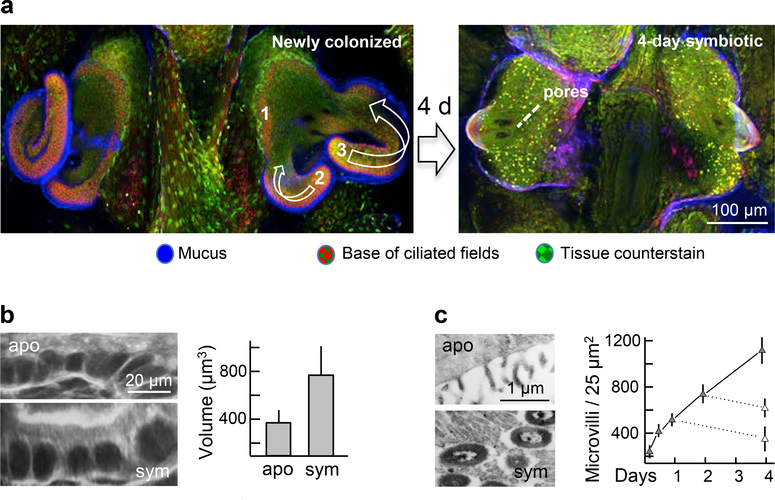
References
-
- Boettcher KJ, Ruby EG, McFall-Ngai MJ. 1996. Bioluminescence in the symbiotic squid Euprymna scolopes is controlled by a daily biological rhythm. J Comp Physiol 179:65–73
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
