

Self-organization of protrusions and polarity during eukaryotic chemotaxis
- PMID: 24998184
- PMCID: PMC4177965
- DOI: 10.1016/j.ceb.2014.06.007
Self-organization of protrusions and polarity during eukaryotic chemotaxis
Abstract
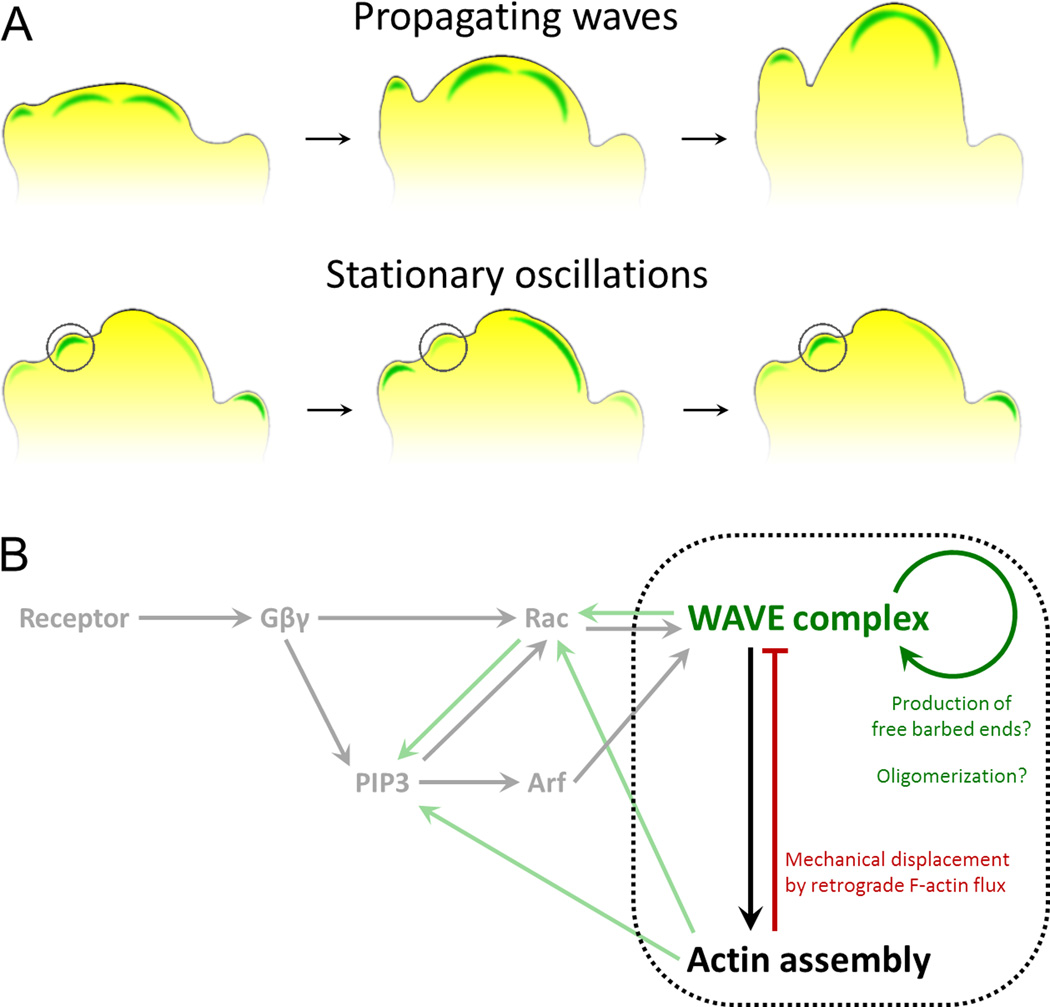
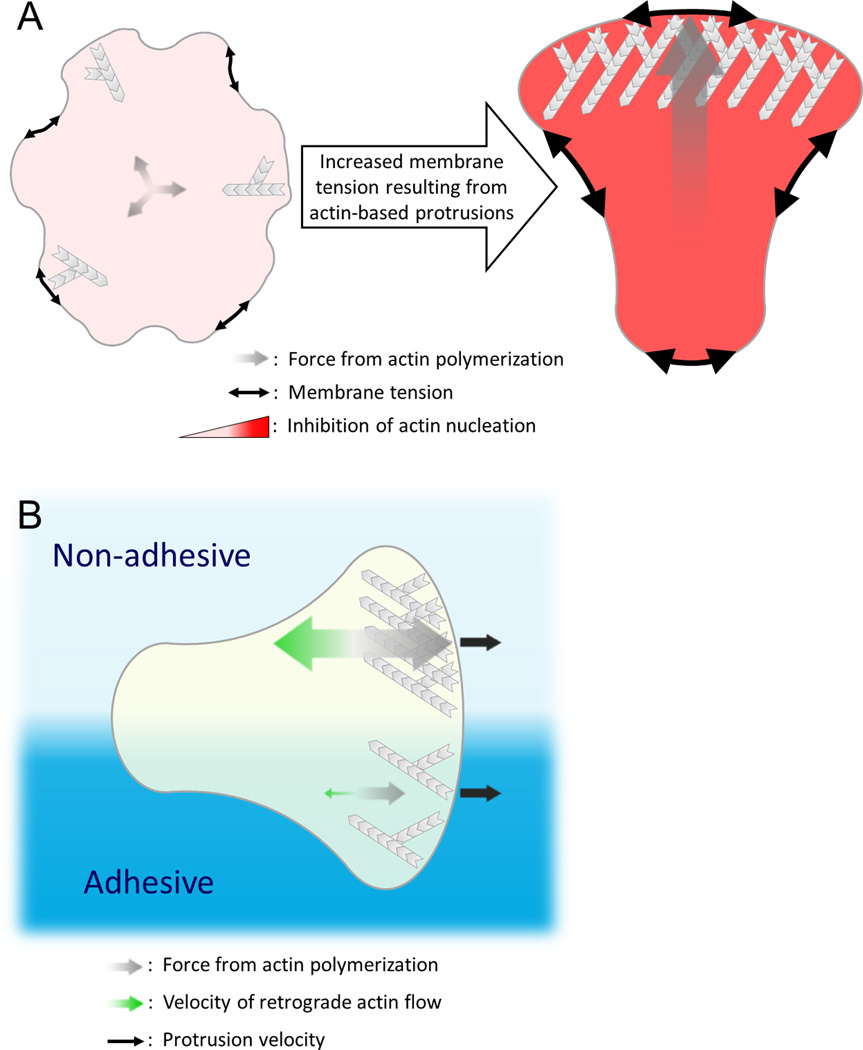
Many eukaryotic cells regulate their polarity and motility in response to external chemical cues. While we know many of the linear connections that link receptors with downstream actin polymerization events, we have a much murkier understanding of the higher order positive and negative feedback loops that organize these processes in space and time. Importantly, physical forces and actin polymerization events do not simply act downstream of chemotactic inputs but are rather involved in a web of reciprocal interactions with signaling components to generate self-organizing pseudopods and cell polarity. Here we focus on recent progress and open questions in the field, including the basic unit of actin organization, how cells regulate the number and speed of protrusions, and 2D versus 3D migration.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
References
-
- De Bruyn PP. The amoeboid movement of the mammalian leukocyte in tissue culture. Anat Rec. 1946;95:177–191. - PubMed
-
-
Killich T, Plath PJ, Wei X, Bultmann H, Rensing L, Vicker MG. The locomotion, shape and pseudopodial dynamics of unstimulated Dictyostelium cells are not random. J Cell Sci. 1993;106(Pt 4):1005–1013. Through careful quantitative measure of cell shape and pseudopod dynamics, this paper provides evidence that the actin cytoskeleton is an oscillatory system more than a decade before fluorescence imaging enabled direct visualization of oscillatory and wavelike actin dynamics.
-
-
- Vicker MG. Reaction-diffusion waves of actin filament polymerization/depolymerization in Dictyostelium pseudopodium extension and cell locomotion. Biophys Chem. 2000;84:87–98. - PubMed
-
- Vicker MG. Eukaryotic cell locomotion depends on the propagation of self-organized reaction-diffusion waves and oscillations of actin filament assembly. Exp Cell Res. 2002;275:54–66. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
