

Review
doi: 10.1038/nrmicro3299.
Epub 2014 Jul 7.
How sisters grow apart: mycobacterial growth and division
Affiliations
- PMID: 24998739
- PMCID: PMC6556109
- DOI: 10.1038/nrmicro3299
Item in Clipboard
Review
How sisters grow apart: mycobacterial growth and division
Nat Rev Microbiol.
2014 Aug.
Abstract
Mycobacterium tuberculosis, which is the aetiological agent of tuberculosis, owes much of its success as a pathogen to its unique cell wall and unusual mechanism of growth, which facilitate its adaptation to the human host and could have a role in clinical latency. Asymmetric growth and division increase population heterogeneity, which may promote antibiotic tolerance and the fitness of single cells. In this Review, we describe the unusual mechanisms of mycobacterial growth, cell wall biogenesis and division, and discuss how these processes might affect the survival of M. tuberculosis in vivo and contribute to the persistence of infection.
Figures
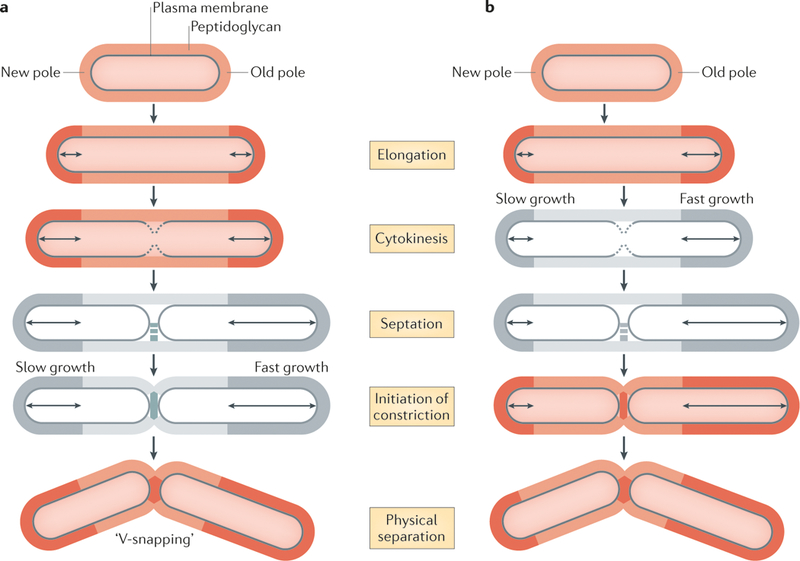
Mycobacterial cells elongate by incorporating new cell wall material (dark orange and dark grey) at their cell poles, and all mother cells contain an old pole (which was inherited from the previous cell division) and a new pole (which was synthesized during the previous cell division). The process of cell division can be subdivided into several stages: synthesis of the plasma membrane roughly at mid-cell during cytokinesis; synthesis of a peptidoglycan septum at mid-cell (which is known as septation); constriction of the cell wall; and hydrolysis of the septum as the two cells physically separate. Cell separation in mycobacteria is characterized by a ‘v-snapping’ phenotype. Two models have been proposed to explain how daughter cells of different sizes are generated, and the models differ in terms of their definitions of cell division. a | In the first model, in which cytokinesis is used to define cell division, growth at both cell poles is symmetric until cytokinesis. From cytokinesis until the cells physically separate, growth occurs predominantly at the old pole, which generates two daughter cells of unequal sizes. In this model, septation and the initiation of constriction were not specifically measured (cells shown in grey). b | In the second model, in which the initiation of constriction during the physical separation of daughter cells is used to define cell division, the poles elongate at different rates throughout the cell cycle, and the old pole elongates faster than the new pole at all stages of the process. This model did not specifically measure cytokinesis or septation (cells shown in grey). Despite discrepancies in the timing of asymmetric growth, both models show that mycobacterial growth generates daughter cells of unequal sizes.
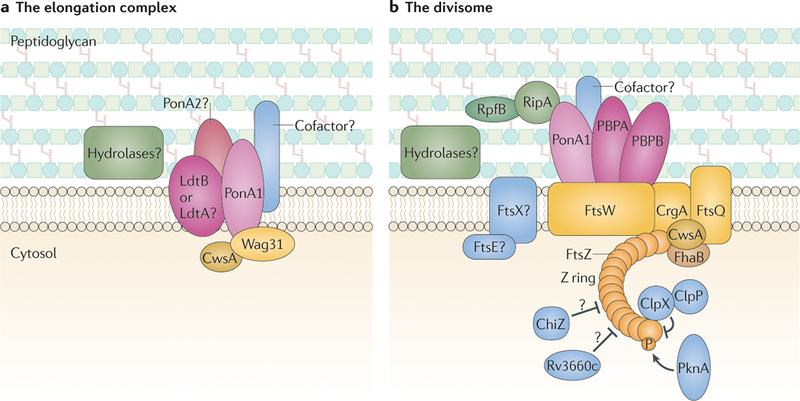
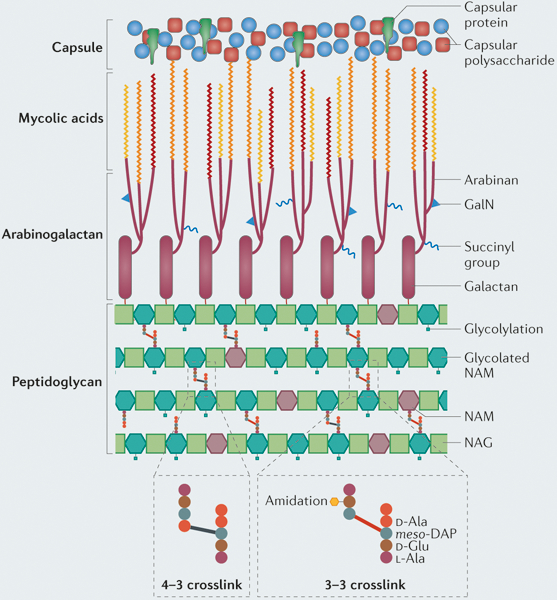
Large macromolecular complexes that are comprised of peptidoglycan synthases (pink), peptidoglycan hydrolases (green), structural proteins (yellow) and regulatory factors (blue) coordinate cell elongation and cell division. a | The multiprotein elongation complex drives polar elongation and is anchored at the poles by Wag31, which is stabilized by cell wall synthesis protein A (CwsA). The peptidoglycan synthase PonAl incorporates new peptidoglycan subunits into the spaces that are created by the hydrolases. PonA2 and the non-classical transpeptidases LdtA and/or LdtB might coordinate peptidoglycan synthesis with PonAl, which would generate the two different types of crosslinks (4–3 and 3–3 crosslinks) that are observed in mycobacterial peptidoglycan. Protein cofactors may coordinate this activity or activate the peptidoglycan synthases. Peptidoglycan hydrolases are required to open the mesh-like peptidoglycan wall for the insertion of new material; however, the identity of these hydrolases is currently unclear. b | Another large macromolecular machine, known as the divisome, is responsible for cell division. Septation is initiated by polymerization of FtsZ, and the subcellular placement of the Z ring is regulated by several factors, including ClpX, ChiZ and Rv3660c, as well as phosphorylation of FtsZ by PknA. FhaB stabilizes the Z ring, and then the structural proteins FtsW, FtsQ, CrgA and CwsA assemble to form the divisome. The peptidoglycan synthases penicillin-binding protein B (PBPB), PBPA and PonAl synthesize the septum, and the hydrolases RipA and RpfB (and possibly other hydrolases) cleave the septum to separate the daughter cells. Septal hydrolysis might also be facilitated by FtsE and FtsX, but this remains to be confirmed.
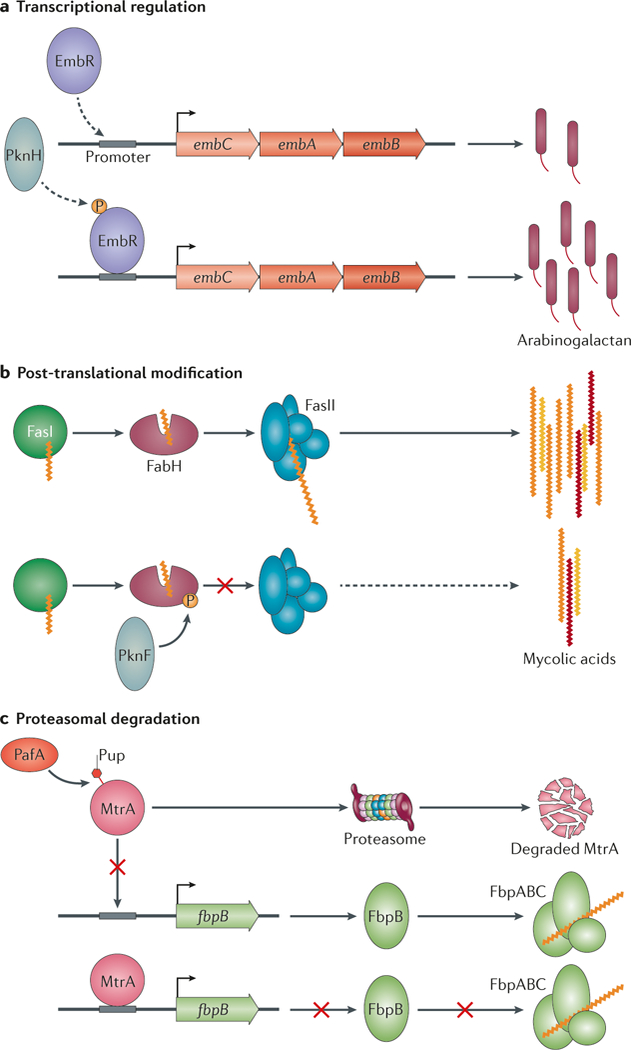
Mycobacteria use several mechanisms to regulate cell wall synthesis and cell division. a | At the transcriptional level, the transcription factor EmbR activates the expression of the embCAB operon, which encodes EmbA, EmbB and EmbC. The two arabinosyltransferases EmbA and EmbB are involved in arabinogalactan synthesis and increase the production of arabinogalactan. Phosphorylation of EmbR by the kinase PknH stimulates promoter binding, which leads to an increase in arabinogalactan synthesis. b | Post-translational modifications of cell wall precursor synthases, such as the phosphorylation of FabH, also occurs. FabH shuttles mycolic acids from FASI to FASII for acyl chain extension, but its activity is inhibited by PknF-mediated phosphorylation, which thereby decreases the production of mycolic acids. c | Mycobacteria encode a unique pupylation system that targets proteins for proteasomal degradation, including proteins that are involved in cell wall synthesis. For example, the response regulator MtrA is pupylated by proteasome accessory factor A (PafA), which promotes its proteasomal degradation. This relieves MtrA-mediated repression of fbpB (which encodes a member of the mycolic acid transferase complex FbpABC, thereby increasing the incorporation of mycolic acids into the cell wall.
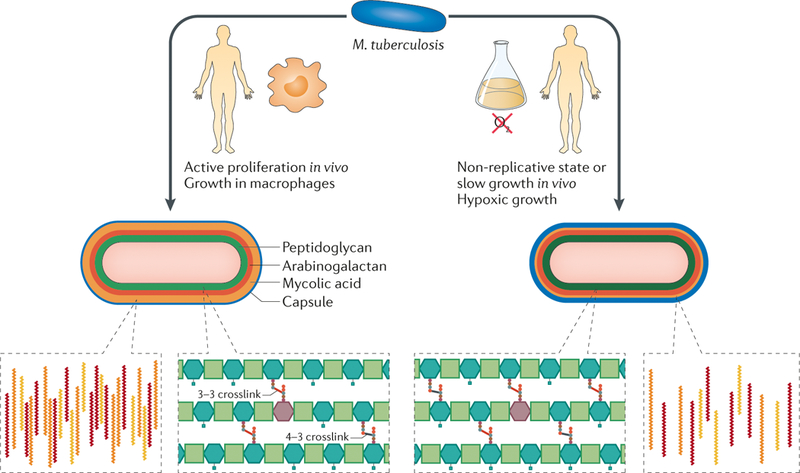
During growth in vivo, substantial changes occur in the cell wall of Mycobacterium tuberculosis. Active proliferation during infection andthe use of host carbon sources during growth in macrophages lead to an increase in the production of mycolic acids and cell wall lipids, such as phthiocerol dimycocerosate (PDIM), which may promote survival in the host environment. In addition, during growth in vivo, peptidoglycan is remodelled and the crucial 3–3 crosslinks that promote chronic infection and antibiotic resistance are maintained. During slow growth and in non-replicative states, the cell wall becomes thicker, probably owing to an increase in the synthesis of multiple cell wall layers. During slow growth and non-replicative states, as well as in hypoxic environments, M. tuberculosis downregulates the abundance of immunostimulatory mycolic acids to potentially avoid immune recognition, and crosslinking of peptidoglycan may be increased to promote cell wall rigidity and bacterial survival under stress. These cell wall changes, combined with the inherent plasticity of cell wall synthesis, generate a population of unique single cells that may have variable fitness during infection.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
