

TRIM68 negatively regulates IFN-β production by degrading TRK fused gene, a novel driver of IFN-β downstream of anti-viral detection systems
- PMID: 24999993
- PMCID: PMC4084880
- DOI: 10.1371/journal.pone.0101503
TRIM68 negatively regulates IFN-β production by degrading TRK fused gene, a novel driver of IFN-β downstream of anti-viral detection systems
Erratum in
-
Correction: TRIM68 Negatively Regulates IFN-β Production by Degrading TRK Fused Gene, a Novel Driver of IFN-β Downstream of Anti-Viral Detection Systems.PLoS One. 2015 Feb 9;10(2):e0117957. doi: 10.1371/journal.pone.0117957. eCollection 2015. PLoS One. 2015. PMID: 25664835 Free PMC article.
Abstract
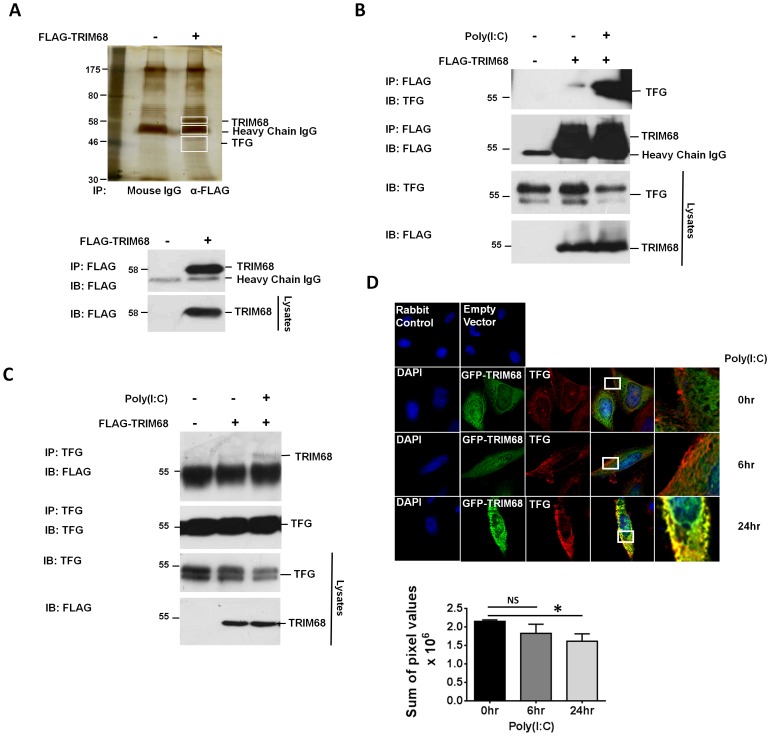
In recent years members of the tripartite motif-containing (TRIM) family of E3 ubiquitin ligases have been shown to both positively and negatively regulate viral defence and as such are emerging as compelling targets for modulating the anti-viral immune response. In this study we identify TRIM68, a close homologue of TRIM21, as a novel regulator of Toll-like receptor (TLR)- and RIG-I-like receptor (RLR)-driven type I IFN production. Proteomic analysis of TRIM68-containing complexes identified TRK-fused gene (TFG) as a potential TRIM68 target. Overexpression of TRIM68 and TFG confirmed their ability to associate, with TLR3 stimulation appearing to enhance the interaction. TFG is a known activator of NF-κB via its ability to interact with inhibitor of NF-κB kinase subunit gamma (IKK-γ) and TRAF family member-associated NF-κB activator (TANK). Our data identifies a novel role for TFG as a positive regulator of type I IFN production and suggests that TRIM68 targets TFG for lysosomal degradation, thus turning off TFG-mediated IFN-β production. Knockdown of TRIM68 in primary human monocytes resulted in enhanced levels of type I IFN and TFG following poly(I:C) treatment. Thus TRIM68 targets TFG, a novel regulator of IFN production, and in doing so turns off and limits type I IFN production in response to anti-viral detection systems.
Conflict of interest statement
Figures



References
-
- Yang B, Wang J, Wang Y, Zhou H, Wu X, et al. (2013) Novel Function of Trim44 Promotes an Antiviral Response by Stabilizing VISA. The Journal of Immunology 190: 3613–3619. - PubMed
-
- Noguchi K, Okumura F, Takahashi N, Kataoka A, Kamiyama T, et al. (2011) TRIM40 promotes neddylation of IKKγ and is downregulated in gastrointestinal cancers. Carcinogenesis 32: 995–1004. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
