

Potential role for HIV-specific CD38-/HLA-DR+ CD8+ T cells in viral suppression and cytotoxicity in HIV controllers
- PMID: 25000587
- PMCID: PMC4084978
- DOI: 10.1371/journal.pone.0101920
Potential role for HIV-specific CD38-/HLA-DR+ CD8+ T cells in viral suppression and cytotoxicity in HIV controllers
Abstract
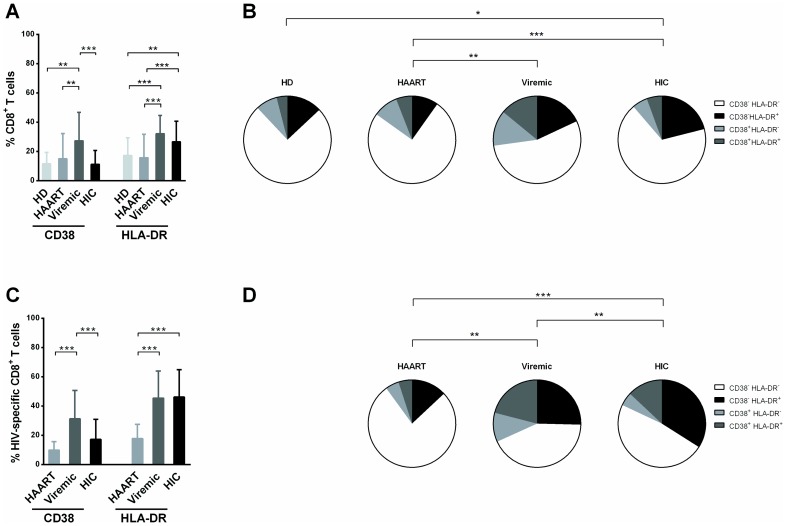
Background: HIV controllers (HIC) are rare HIV-1-infected patients who exhibit spontaneous viral control. HIC have high frequency of CD38-/HLA-DR+ HIV-specific CD8+ T cells. Here we examined the role of this subset in HIC status.
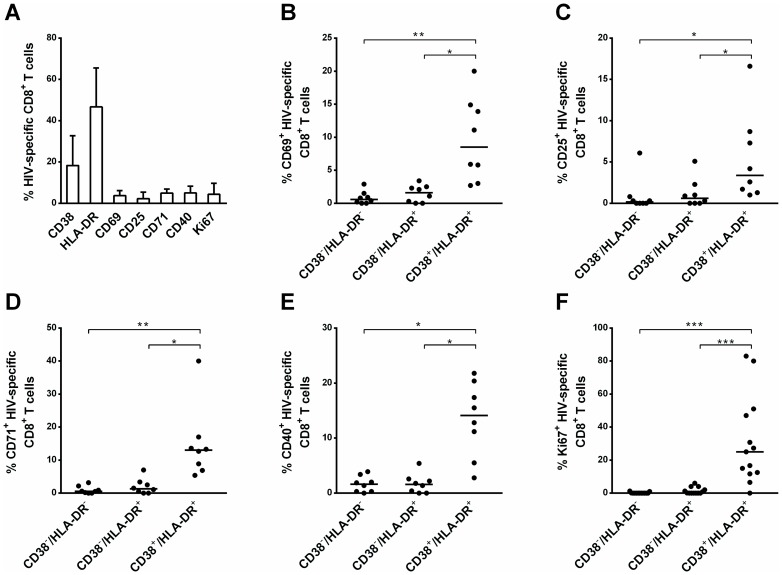
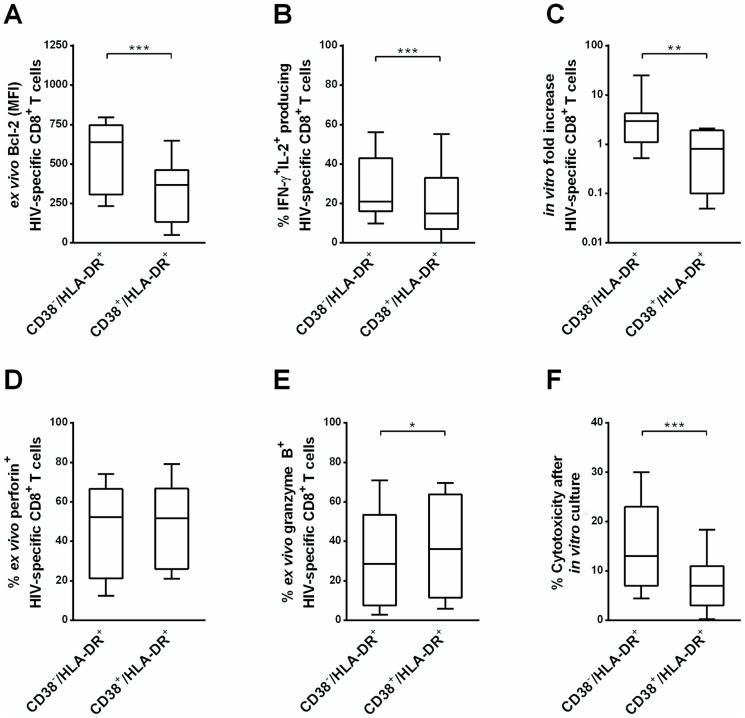
Materials and methods: We compared CD38-/HLA-DR+ CD8+ T cells with the classical CD38+/HLA-DR+ activated phenotype in terms of 1) their activation status, reflected by CD69, CD25, CD71, CD40 and Ki67 expression, 2) functional parameters: Bcl-2 expression, proliferative capacity, and IFN-γ and IL-2 production, and 3) cytotoxic activity. We also investigated how this particular profile is generated.
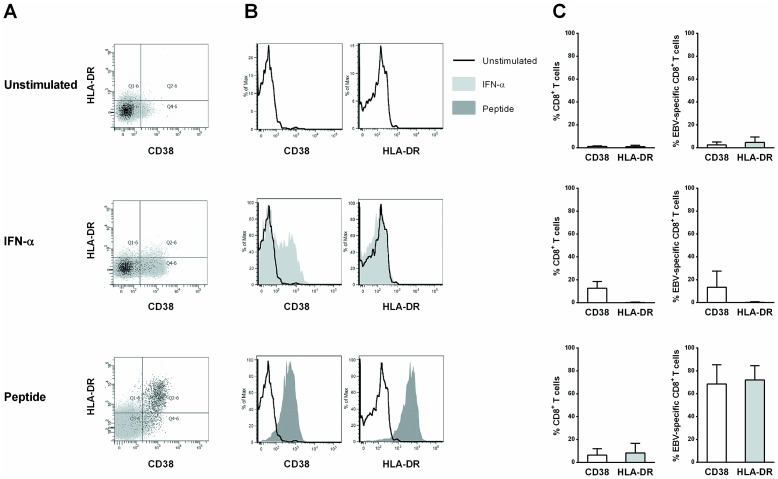
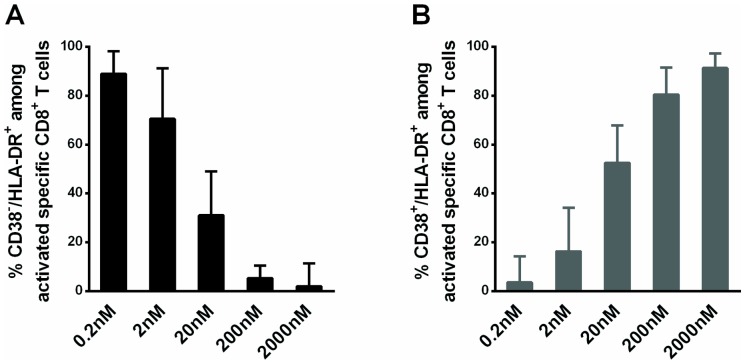
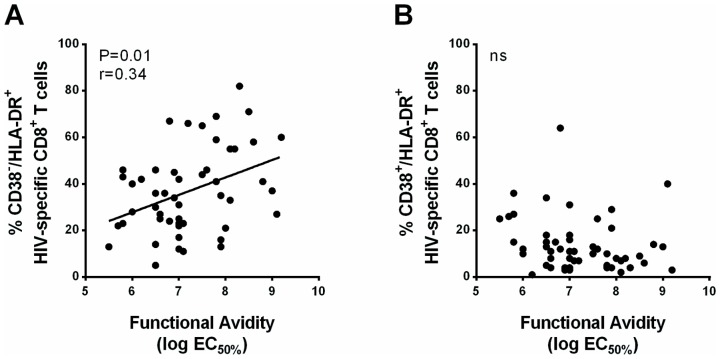
Results: Compared to CD38+/HLA-DR+ cells, CD38-/HLA-DR+ cells exhibited lower expression of several activation markers, better survival capacity (Bcl-2 MFI, 367 [134-462] vs 638 [307-747], P = 0.001), higher frequency of polyfunctional cells (15% [7%-33%] vs 21% [16%-43%], P = 0.0003), greater proliferative capacity (0-fold [0-2] vs 3-fold [2]-[11], P = 0.007), and higher cytotoxicity in vitro (7% [3%-11%] vs 13% [6%-22%], P = 0.02). The CD38-/HLA-DR+ profile was preferentially generated in response to low viral antigen concentrations.
Conclusions: These data highlight the role of CD38-/HLA-DR+ HIV-specific CD8+ T cell cytotoxicity in HIC status and provide insights into the mechanism by which they are generated. Induction of this protective CD8+ subset may be important for vaccine strategies.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
