

Memory CD8(+) T cells use cell-intrinsic lipolysis to support the metabolic programming necessary for development
- PMID: 25001241
- PMCID: PMC4120664
- DOI: 10.1016/j.immuni.2014.06.005
Memory CD8(+) T cells use cell-intrinsic lipolysis to support the metabolic programming necessary for development
Erratum in
-
Memory CD8+ T Cells Use Cell-Intrinsic Lipolysis to Support the Metabolic Programming Necessary for Development.Immunity. 2018 Aug 21;49(2):375-376. doi: 10.1016/j.immuni.2018.07.018. Immunity. 2018. PMID: 30134202 Free PMC article. No abstract available.
Abstract
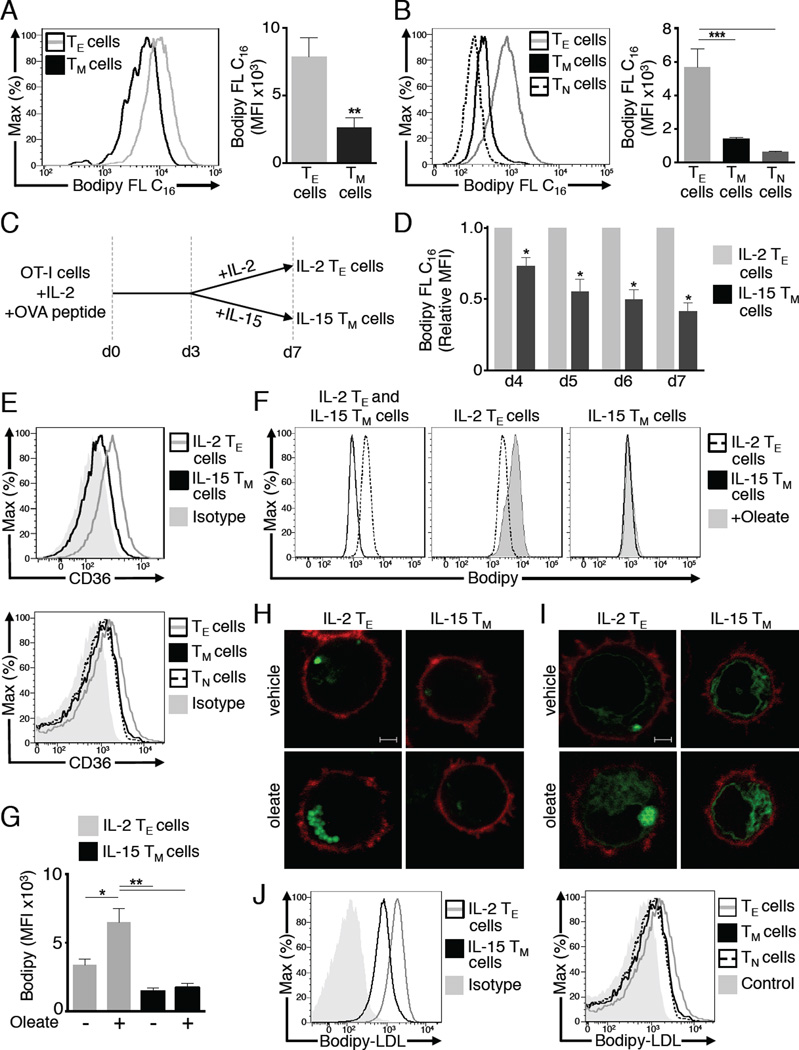
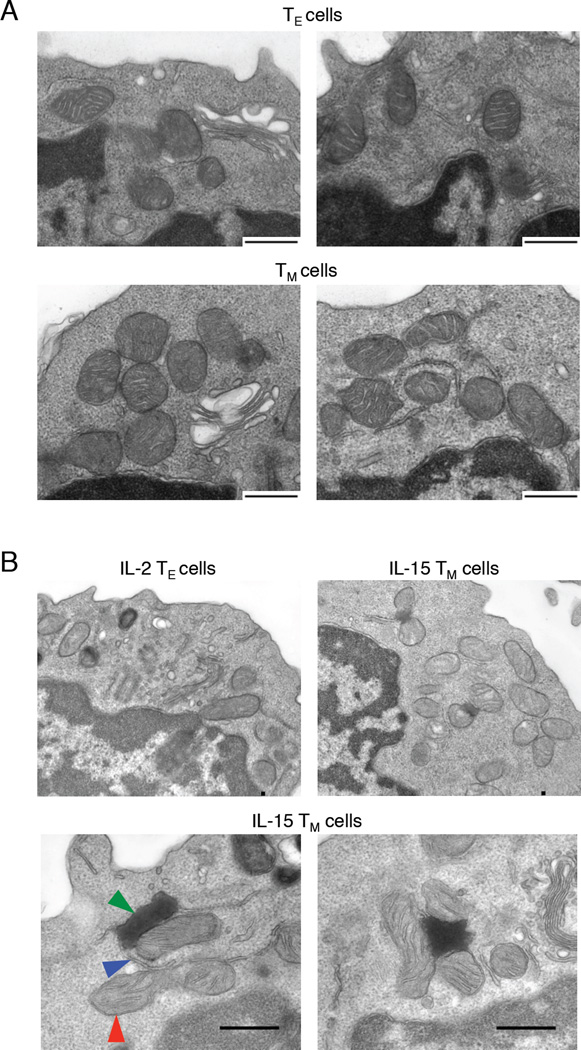
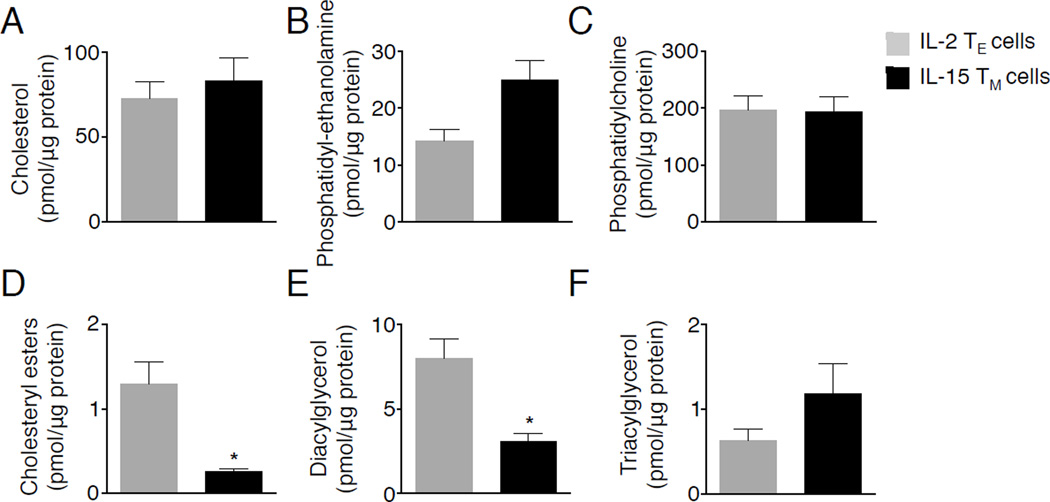
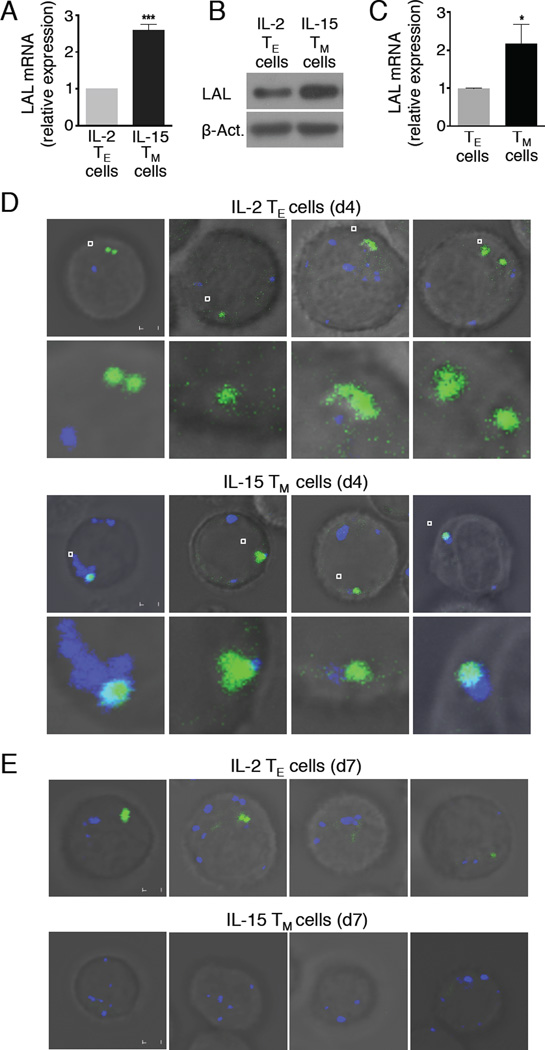
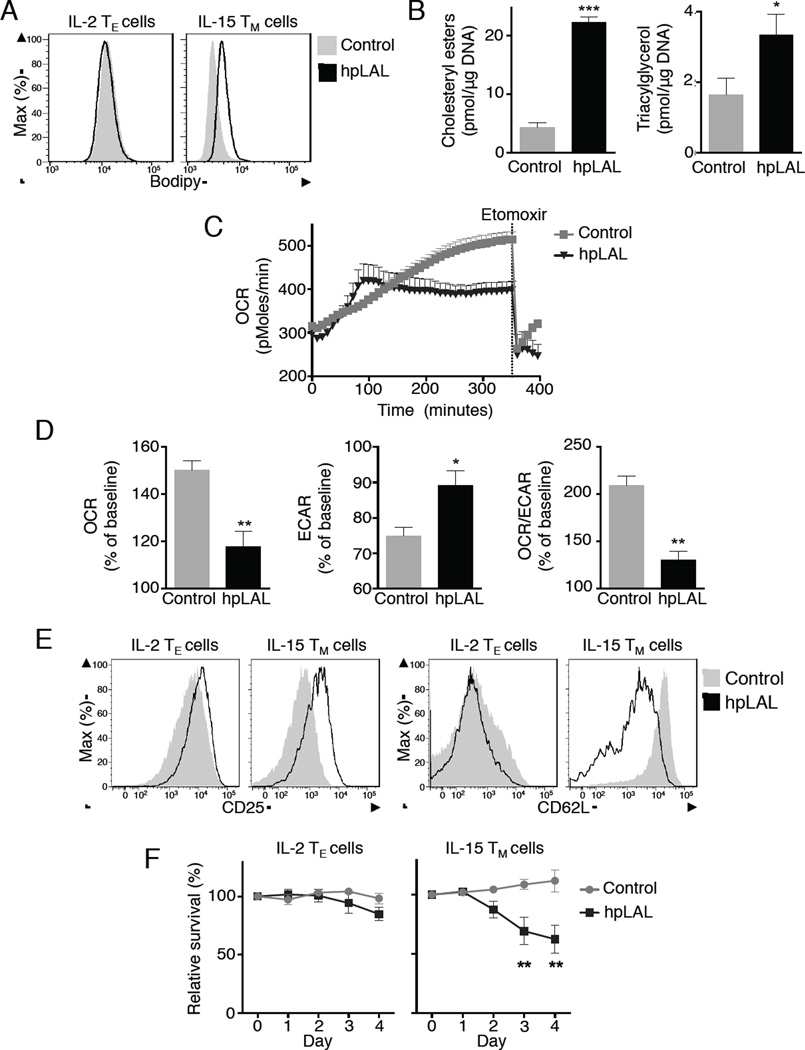
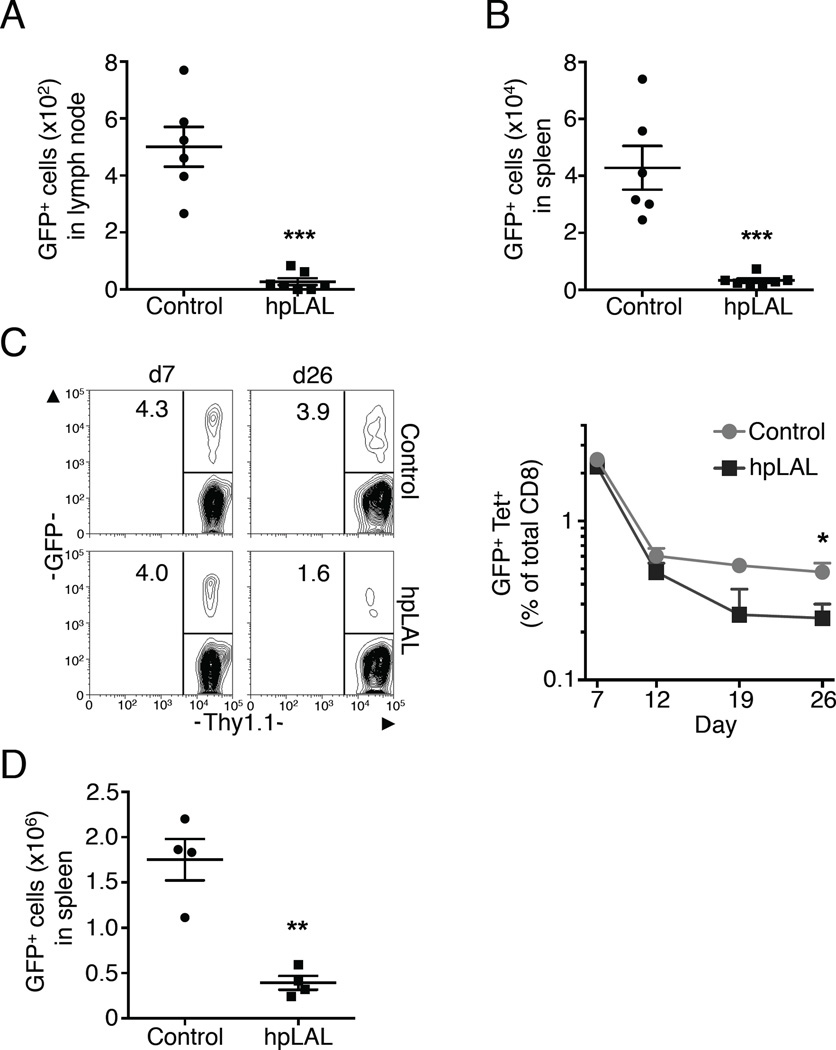
Generation of CD8(+) memory T cells requires metabolic reprogramming that is characterized by enhanced mitochondrial fatty-acid oxidation (FAO). However, where the fatty acids (FA) that fuel this process come from remains unclear. While CD8(+) memory T cells engage FAO to a greater extent, we found that they acquired substantially fewer long-chain FA from their external environment than CD8(+) effector T (Teff) cells. Rather than using extracellular FA directly, memory T cells used extracellular glucose to support FAO and oxidative phosphorylation (OXPHOS), suggesting that lipids must be synthesized to generate the substrates needed for FAO. We have demonstrated that memory T cells rely on cell intrinsic expression of the lysosomal hydrolase LAL (lysosomal acid lipase) to mobilize FA for FAO and memory T cell development. Our observations link LAL to metabolic reprogramming in lymphocytes and show that cell intrinsic lipolysis is deterministic for memory T cell fate.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures

Comment in
-
T cell memory: Metabolic self-reliance.Nat Rev Immunol. 2014 Aug;14(8):518. doi: 10.1038/nri3721. Epub 2014 Jul 18. Nat Rev Immunol. 2014. PMID: 25033905 No abstract available.
-
Futility sustains memory T cells.Immunity. 2014 Jul 17;41(1):1-3. doi: 10.1016/j.immuni.2014.06.009. Immunity. 2014. PMID: 25035944 Free PMC article.
References
-
- Bowden KL, Bilbey NJ, Bilawchuk LM, Boadu E, Sidhu R, Ory DS, Du H, Chan T, Francis GA. Lysosomal Acid Lipase Deficiency Impairs Regulation of ABCA1 Gene and Formation of High Density Lipoproteins in Cholesteryl Ester Storage Disease. Journal of Biological Chemistry. 2011;286:30624–30635. - PMC - PubMed
-
- Brasaemle DL. Thematic review series: Adipocyte Biology. The perilipin family of structural lipid droplet proteins: stabilization of lipid droplets and control of lipolysis. Journal of Lipid Research. 2007;48:2547–2559. - PubMed
-
- Buzzai M, Bauer DE, Jones RG, DeBerardinis RJ, Hatzivassiliou G, Elstrom RL, Thompson CB. The glucose dependence of Akt-transformed cells can be reversed by pharmacologic activation of fatty acid [beta]-oxidation. Oncogene. 2005;24:4165–4173. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
