

Sonic hedgehog-expressing cells in the developing limb measure time by an intrinsic cell cycle clock
- PMID: 25001275
- PMCID: PMC4102111
- DOI: 10.1038/ncomms5230
Sonic hedgehog-expressing cells in the developing limb measure time by an intrinsic cell cycle clock
Abstract
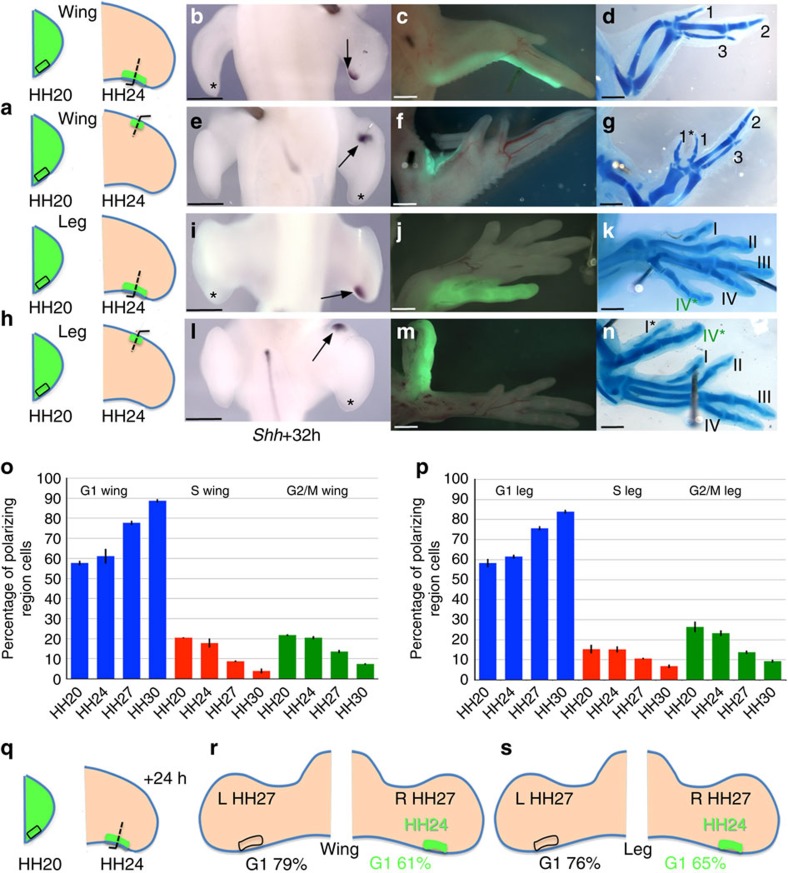
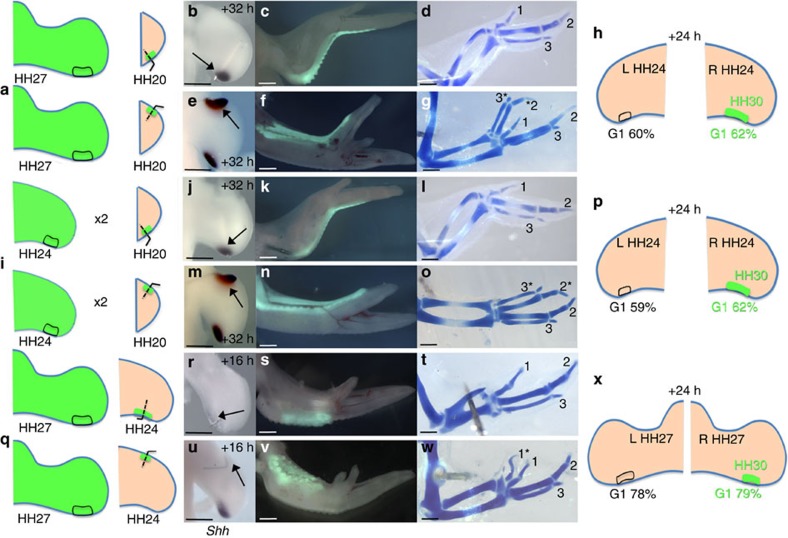
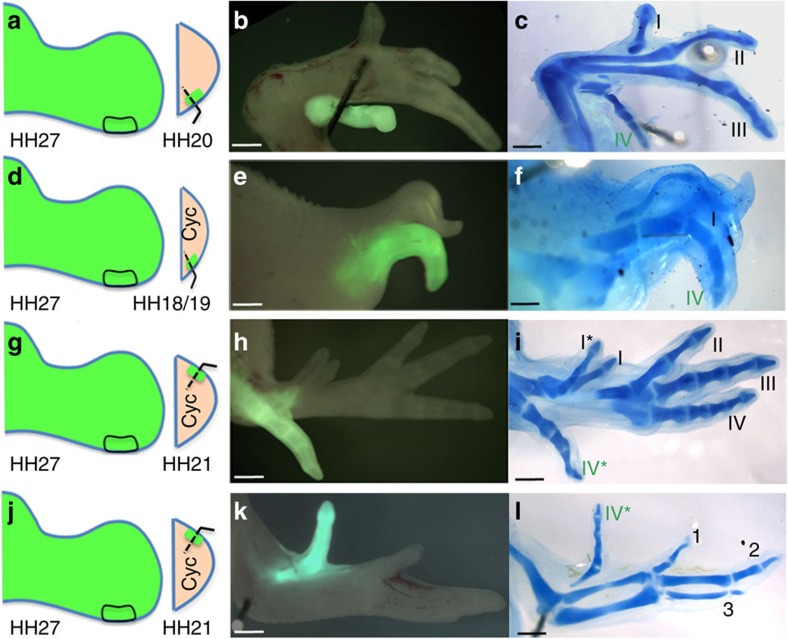
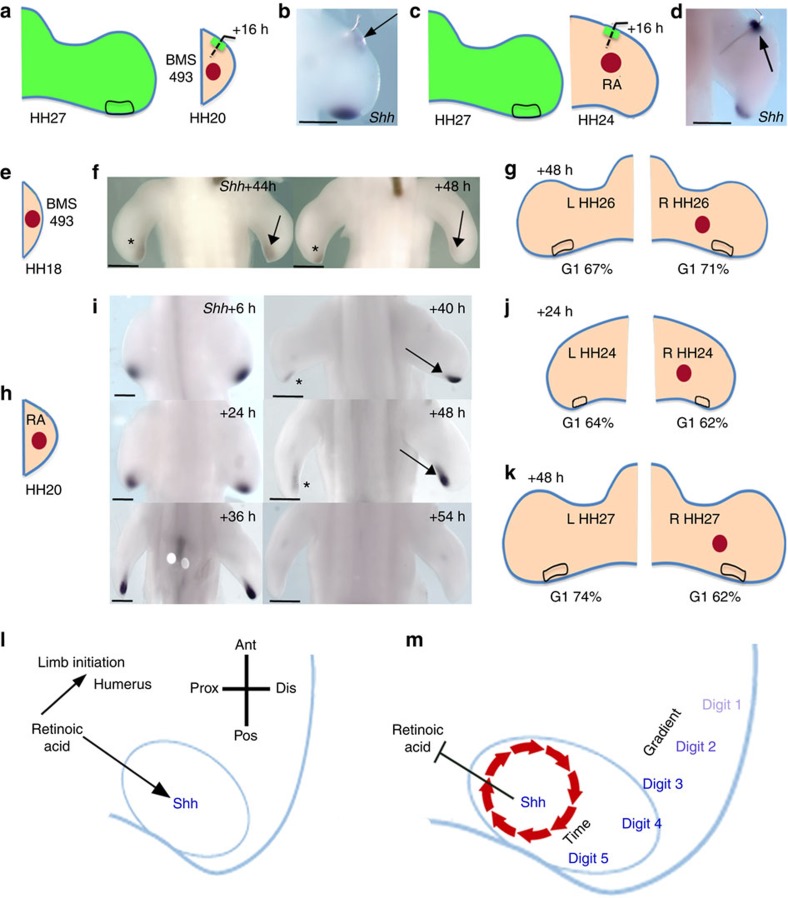
How time is measured is an enduring issue in developmental biology. Classical models of somitogenesis and limb development implicated intrinsic cell cycle clocks, but their existence remains controversial. Here we show that an intrinsic cell cycle clock in polarizing region cells of the chick limb bud times the duration of Sonic hedgehog (Shh) expression, which encodes the morphogen specifying digit pattern across the antero-posterior axis (thumb to little finger). Timing by this clock starts when polarizing region cells fall out of range of retinoic acid signalling. We found that timing of Shh transcription by the cell cycle clock can be reset, thus revealing an embryonic form of self-renewal. In contrast, antero-posterior positional values cannot be reset, suggesting that this may be an important constraint on digit regeneration. Our findings provide the first evidence for an intrinsic cell cycle timer controlling duration and patterning activity of a major embryonic signalling centre.
Figures
Similar articles
-
The Sonic hedgehog gradient in the developing limb.Wiley Interdiscip Rev Dev Biol. 2013 Mar-Apr;2(2):275-90. doi: 10.1002/wdev.70. Epub 2012 May 25. Wiley Interdiscip Rev Dev Biol. 2013. PMID: 24009037
-
How the embryo makes a limb: determination, polarity and identity.J Anat. 2015 Oct;227(4):418-30. doi: 10.1111/joa.12361. Epub 2015 Aug 7. J Anat. 2015. PMID: 26249743 Free PMC article. Review.
-
An autoregulatory cell cycle timer integrates growth and specification in chick wing digit development.Elife. 2019 Sep 23;8:e47625. doi: 10.7554/eLife.47625. Elife. 2019. PMID: 31545166 Free PMC article.
-
Expression of genes encoding bone morphogenetic proteins and sonic hedgehog in talpid (ta3) limb buds: their relationships in the signalling cascade involved in limb patterning.Dev Dyn. 1995 Jun;203(2):187-97. doi: 10.1002/aja.1002030207. Dev Dyn. 1995. PMID: 7655081
-
Gradients of signalling in the developing limb.Curr Opin Cell Biol. 2012 Apr;24(2):181-7. doi: 10.1016/j.ceb.2011.11.005. Epub 2011 Dec 13. Curr Opin Cell Biol. 2012. PMID: 22169676 Review.
Cited by
-
The Hedgehog signalling pathway in bone formation.Int J Oral Sci. 2015 Jun 26;7(2):73-9. doi: 10.1038/ijos.2015.14. Int J Oral Sci. 2015. PMID: 26023726 Free PMC article. Review.
-
An intrinsic timer specifies distal structures of the vertebrate limb.Nat Commun. 2015 Sep 18;6:8108. doi: 10.1038/ncomms9108. Nat Commun. 2015. PMID: 26381580 Free PMC article.
-
Retinoic acid influences the timing and scaling of avian wing development.Cell Rep. 2022 Jan 25;38(4):110288. doi: 10.1016/j.celrep.2021.110288. Cell Rep. 2022. PMID: 35081337 Free PMC article.
-
Tissue tectonics and the multi-scale regulation of developmental timing.Interface Focus. 2021 Apr 16;11(3):20200057. doi: 10.1098/rsfs.2020.0057. eCollection 2021 Jun 6. Interface Focus. 2021. PMID: 34055304 Free PMC article. Review.
-
The PROTAC selectively degrading Bcl-xL represents a novel Hedgehog pathway inhibitor with capacity of combating resistance to Smoothened inhibitors while sparing bone growth.Theranostics. 2022 Oct 24;12(17):7476-7490. doi: 10.7150/thno.75421. eCollection 2022. Theranostics. 2022. PMID: 36438482 Free PMC article.
References
-
- Primmett D. R., Norris W. E., Carlson G. J., Keynes R. J. & Stern C. D. Periodic segmental anomalies induced by heat shock in the chick embryo are associated with the cell cycle. Development 105, 119–130 (1989). - PubMed
-
- Primmett D. R., Stern C. D. & Keynes R. J. Heat shock causes repeated segmental anomalies in the chick embryo. Development 104, 331–339 (1988). - PubMed
-
- Duboule D. Temporal colinearity and the phylotypic progression: a basis for the stability of a vertebrate Bauplan and the evolution of morphologies through heterochrony. Development 135–142 (1994). - PubMed
-
- Summerbell D., Lewis J. H. & Wolpert L. Positional information in chick limb morphogenesis. Nature 244, 492–496 (1973). - PubMed
-
- Lewis J. H. Fate maps and the pattern of cell division: a calculation for the chick wing-bud. J. Embryol. Exp. Morphol. 33, 419–434 (1975). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
