

Proteomic mapping of the human mitochondrial intermembrane space in live cells via ratiometric APEX tagging
- PMID: 25002142
- PMCID: PMC4743503
- DOI: 10.1016/j.molcel.2014.06.003
Proteomic mapping of the human mitochondrial intermembrane space in live cells via ratiometric APEX tagging
Abstract
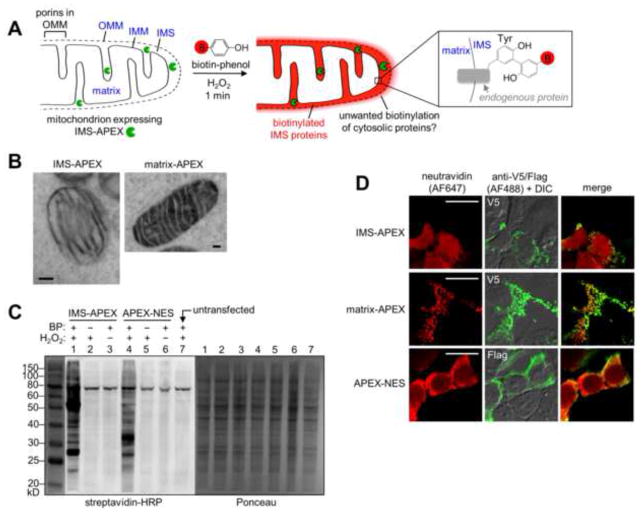
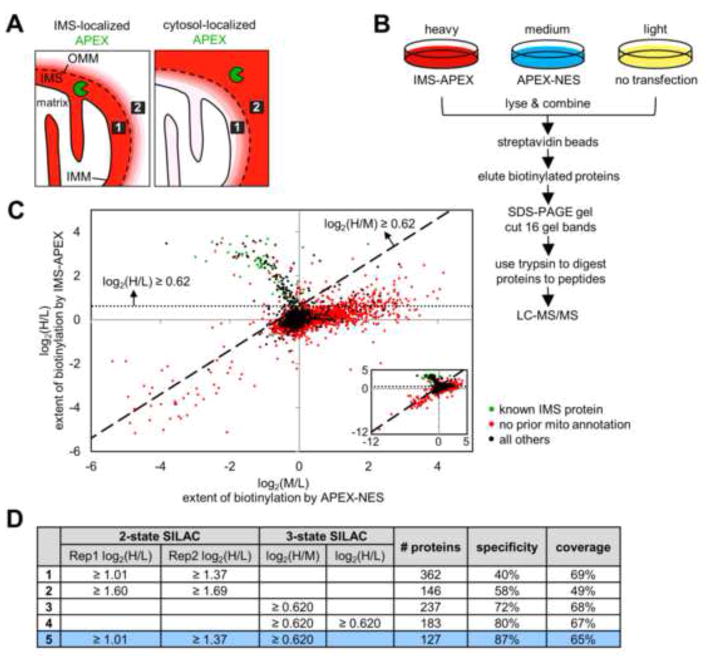
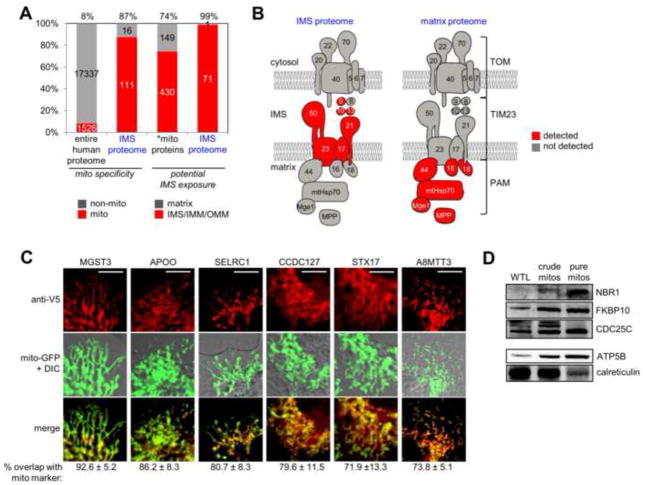
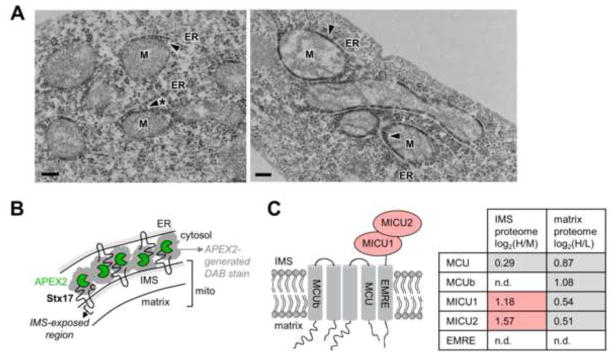
Obtaining complete protein inventories for subcellular regions is a challenge that often limits our understanding of cellular function, especially for regions that are impossible to purify and are therefore inaccessible to traditional proteomic analysis. We recently developed a method to map proteomes in living cells with an engineered peroxidase (APEX) that bypasses the need for organellar purification when applied to membrane-bound compartments; however, it was insufficiently specific when applied to unbounded regions that allow APEX-generated radicals to escape. Here, we combine APEX technology with a SILAC-based ratiometric tagging strategy to substantially reduce unwanted background and achieve nanometer spatial resolution. This is applied to map the proteome of the mitochondrial intermembrane space (IMS), which can freely exchange small molecules with the cytosol. Our IMS proteome of 127 proteins has >94% specificity and includes nine newly discovered mitochondrial proteins. This approach will enable scientists to map proteomes of cellular regions that were previously inaccessible.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- del Arco A, Satrústegui J. Identification of a Novel Human Subfamily of Mitochondrial Carriers with Calcium-binding Domains. J Biol Chem. 2004;279:24701–24713. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
