

Phosphorylation and degradation of tomosyn-2 de-represses insulin secretion
- PMID: 25002582
- PMCID: PMC4155689
- DOI: 10.1074/jbc.M114.575985
Phosphorylation and degradation of tomosyn-2 de-represses insulin secretion
Abstract
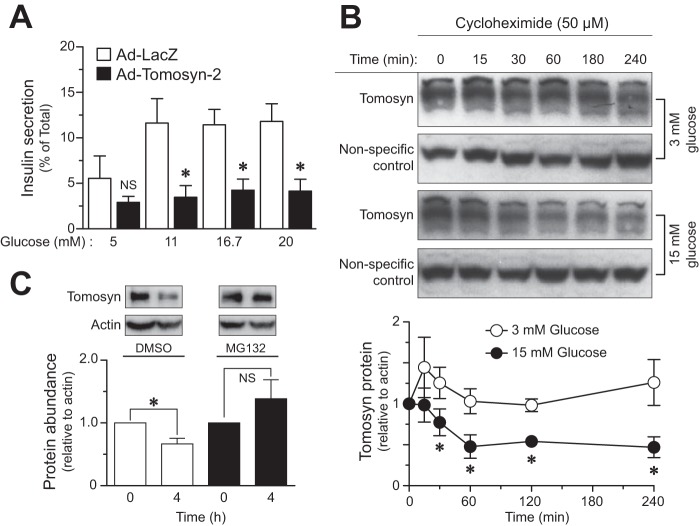
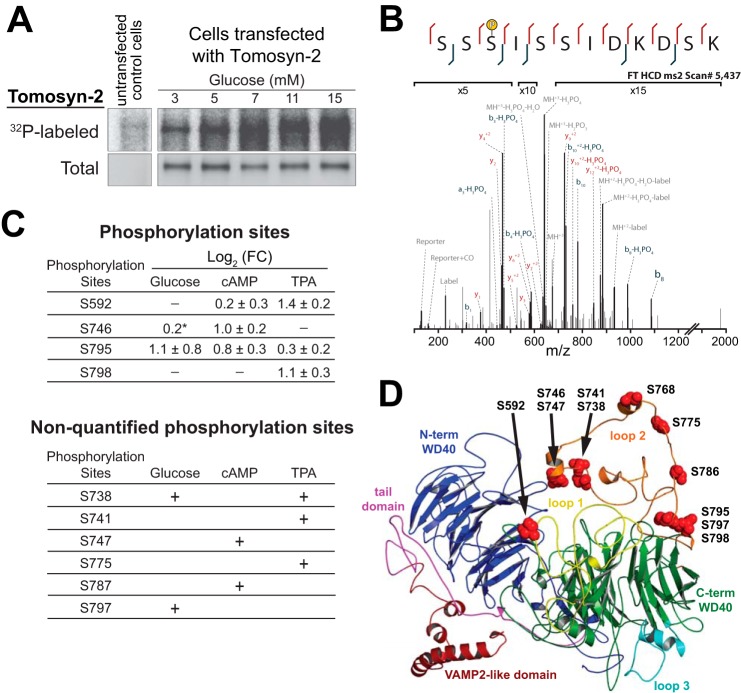
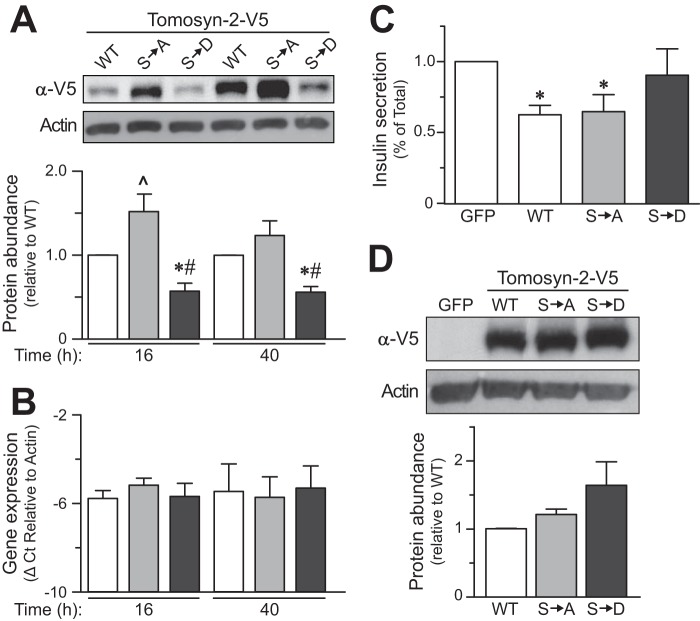
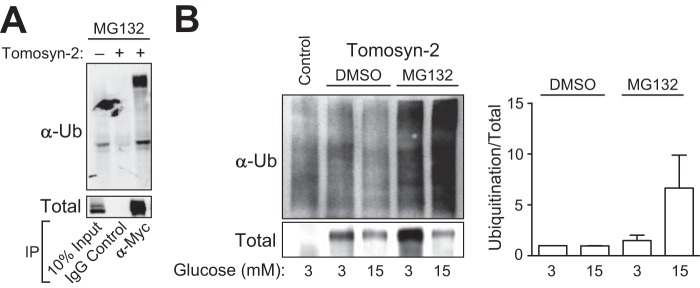
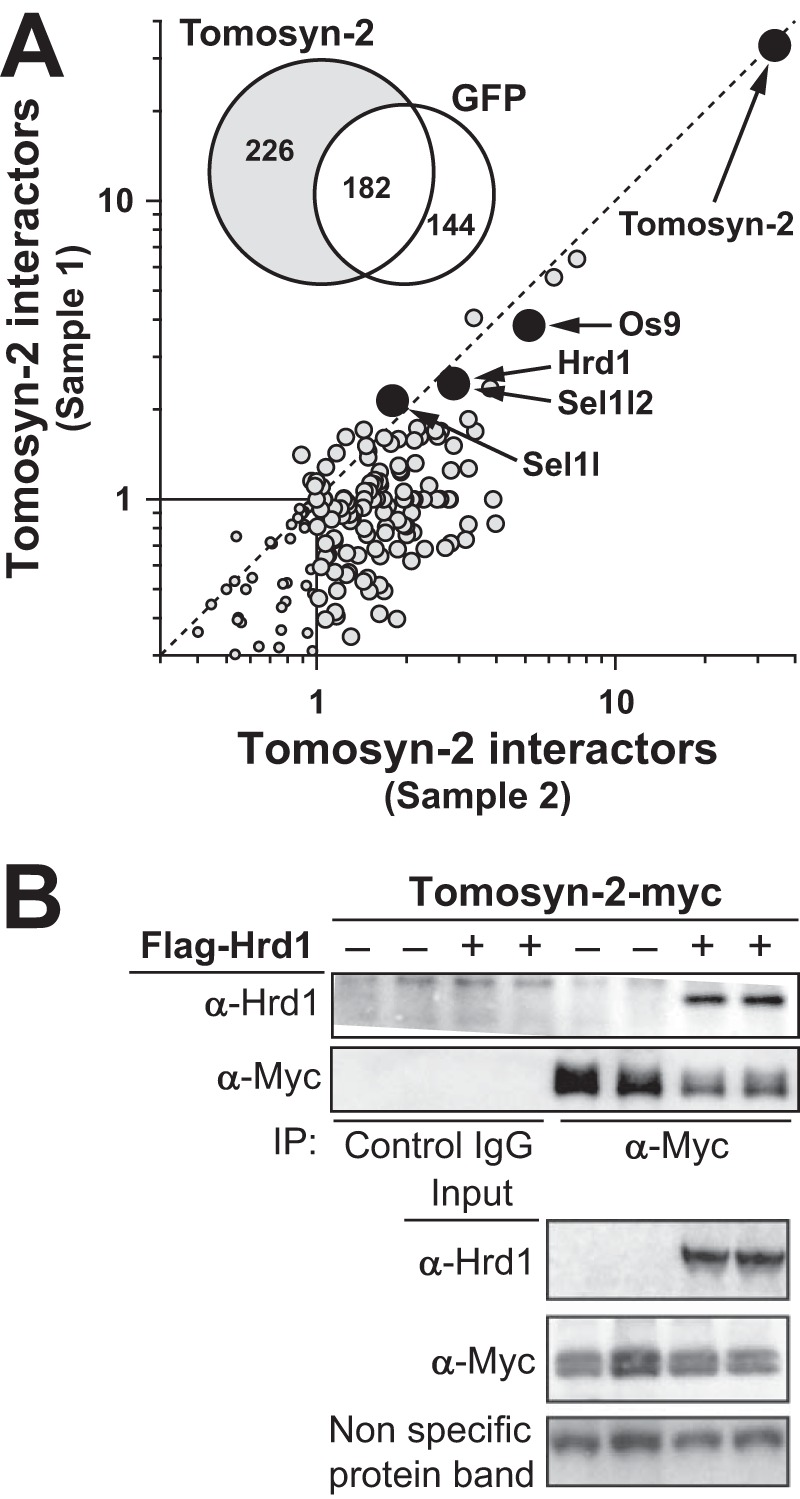
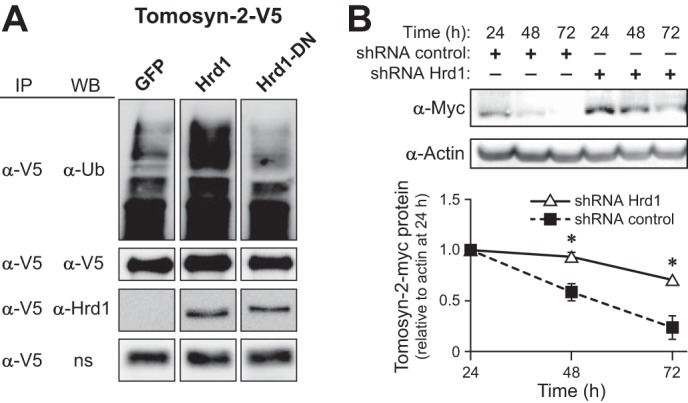
The abundance and functional activity of proteins involved in the formation of the SNARE complex are tightly regulated for efficient exocytosis. Tomosyn proteins are negative regulators of exocytosis. Tomosyn causes an attenuation of insulin secretion by limiting the formation of the SNARE complex. We hypothesized that glucose-dependent stimulation of insulin secretion from β-cells must involve reversing the inhibitory action of tomosyn. Here, we show that glucose increases tomosyn protein turnover. Within 1 h of exposure to 15 mM glucose, ~50% of tomosyn was degraded. The degradation of tomosyn in response to high glucose was blocked by inhibitors of the proteasomal pathway. Using (32)P labeling and mass spectrometry, we showed that tomosyn-2 is phosphorylated in response to high glucose, phorbol esters, and analogs of cAMP, all key insulin secretagogues. We identified 11 phosphorylation sites in tomosyn-2. Site-directed mutagenesis was used to generate phosphomimetic (Ser → Asp) and loss-of-function (Ser → Ala) mutants. The Ser → Asp mutant had enhanced protein turnover compared with the Ser → Ala mutant and wild type tomosyn-2. Additionally, the Ser → Asp tomosyn-2 mutant was ineffective at inhibiting insulin secretion. Using a proteomic screen for tomosyn-2-binding proteins, we identified Hrd-1, an E3-ubiquitin ligase. We showed that tomosyn-2 ubiquitination is increased by Hrd-1, and knockdown of Hrd-1 by short hairpin RNA resulted in increased abundance in tomosyn-2 protein levels. Taken together, our results reveal a mechanism by which enhanced phosphorylation of a negative regulator of secretion, tomosyn-2, in response to insulin secretagogues targets it to degradation by the Hrd-1 E3-ubiquitin ligase.
Keywords: Diabetes; Insulin; Insulin Secretion; SNARE Proteins; Type 2 Diabetes.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
Similar articles
-
Positional cloning of a type 2 diabetes quantitative trait locus; tomosyn-2, a negative regulator of insulin secretion.PLoS Genet. 2011 Oct;7(10):e1002323. doi: 10.1371/journal.pgen.1002323. Epub 2011 Oct 6. PLoS Genet. 2011. PMID: 21998599 Free PMC article.
-
Tomosyn-1 is involved in a post-docking event required for pancreatic beta-cell exocytosis.J Cell Sci. 2006 Jul 15;119(Pt 14):2912-20. doi: 10.1242/jcs.03037. Epub 2006 Jun 20. J Cell Sci. 2006. PMID: 16787939
-
Genetic ablation of synaptotagmin-9 alters tomosyn-1 function to increase insulin secretion from pancreatic β-cells improving glucose clearance.FASEB J. 2023 Aug;37(8):e23075. doi: 10.1096/fj.202300291RR. FASEB J. 2023. PMID: 37432648 Free PMC article.
-
Tomosyn is a novel Akt substrate mediating insulin-dependent GLUT4 exocytosis.Int J Biochem Cell Biol. 2015 May;62:62-71. doi: 10.1016/j.biocel.2015.02.013. Epub 2015 Feb 25. Int J Biochem Cell Biol. 2015. PMID: 25725259
-
Cell-wide analysis of secretory granule dynamics in three dimensions in living pancreatic beta-cells: evidence against a role for AMPK-dependent phosphorylation of KLC1 at Ser517/Ser520 in glucose-stimulated insulin granule movement.Biochem Soc Trans. 2010 Feb;38(Pt 1):205-8. doi: 10.1042/BST0380205. Biochem Soc Trans. 2010. PMID: 20074060 Review.
Cited by
-
The islet-expressed Lhx1 transcription factor interacts with Islet-1 and contributes to glucose homeostasis.Am J Physiol Endocrinol Metab. 2019 Mar 1;316(3):E397-E409. doi: 10.1152/ajpendo.00235.2018. Epub 2019 Jan 8. Am J Physiol Endocrinol Metab. 2019. PMID: 30620636 Free PMC article.
-
Complement 1q-like-3 protein inhibits insulin secretion from pancreatic β-cells via the cell adhesion G protein-coupled receptor BAI3.J Biol Chem. 2018 Nov 23;293(47):18086-18098. doi: 10.1074/jbc.RA118.005403. Epub 2018 Sep 18. J Biol Chem. 2018. PMID: 30228187 Free PMC article.
-
Ranking factors involved in diabetes remission after bariatric surgery using machine-learning integrating clinical and genomic biomarkers.NPJ Genom Med. 2016 Oct 26;1:16035. doi: 10.1038/npjgenmed.2016.35. eCollection 2016. NPJ Genom Med. 2016. PMID: 29263820 Free PMC article.
-
Endoplasmic Reticulum Protein Quality Control in β Cells.Semin Cell Dev Biol. 2020 Jul;103:59-67. doi: 10.1016/j.semcdb.2020.04.006. Epub 2020 May 8. Semin Cell Dev Biol. 2020. PMID: 32402517 Free PMC article. Review.
-
A gene expression network analysis of the pancreatic islets from lean and obese mice identifies complement 1q like-3 secreted protein as a regulator of β-cell function.Sci Rep. 2019 Jul 12;9(1):10119. doi: 10.1038/s41598-019-46219-3. Sci Rep. 2019. PMID: 31300714 Free PMC article.
References
-
- Eddlestone G. T., Oldham S. B., Lipson L. G., Premdas F. H., Beigelman P. M. (1985) Electrical activity, cAMP concentration, and insulin release in mouse islets of Langerhans. Am. J. Physiol. 248, C145–C153 - PubMed
-
- Rorsman P., Eliasson L., Renström E., Gromada J., Barg S., Göpel S. (2000) The cell physiology of biphasic insulin secretion. News Physiol. Sci. 15, 72–77 - PubMed
-
- Straub S. G., Sharp G. W. (2002) Glucose-stimulated signaling pathways in biphasic insulin secretion. Diabetes Metab. Res. Rev. 18, 451–463 - PubMed
-
- Sudhof T. C. (2004) The synaptic vesicle cycle. Annu. Rev. Neurosci. 27, 509–547 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
