

Phosphatase 1 nuclear targeting subunit is an essential regulator of M-phase entry, maintenance, and exit
- PMID: 25002584
- PMCID: PMC4156095
- DOI: 10.1074/jbc.M114.572149
Phosphatase 1 nuclear targeting subunit is an essential regulator of M-phase entry, maintenance, and exit
Abstract
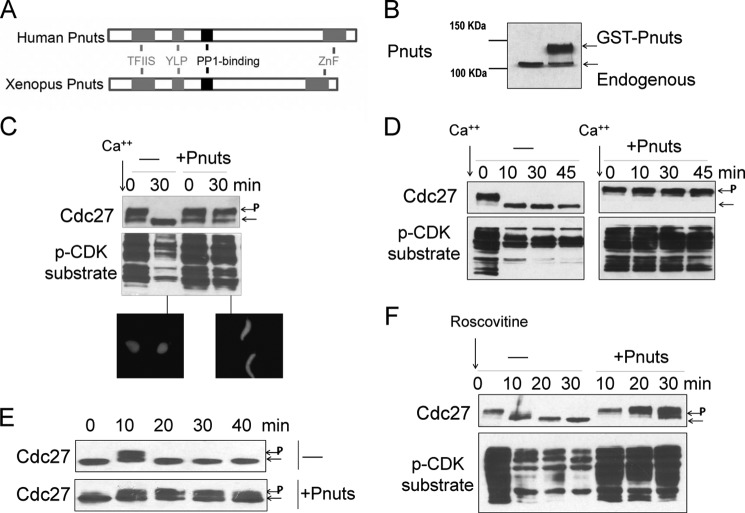
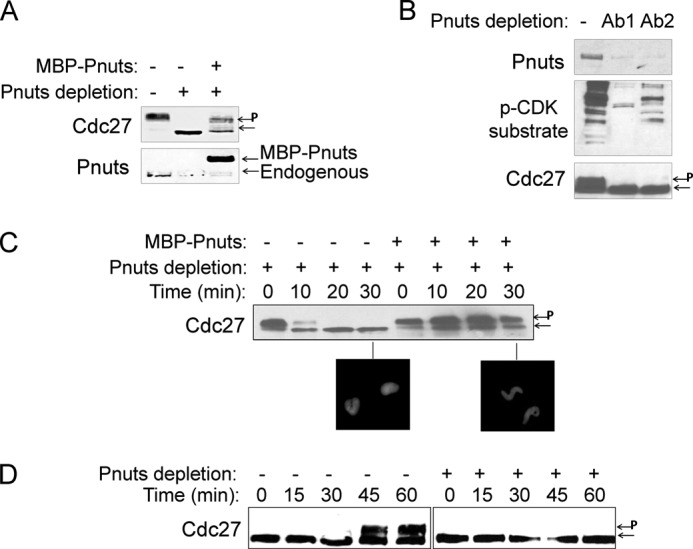
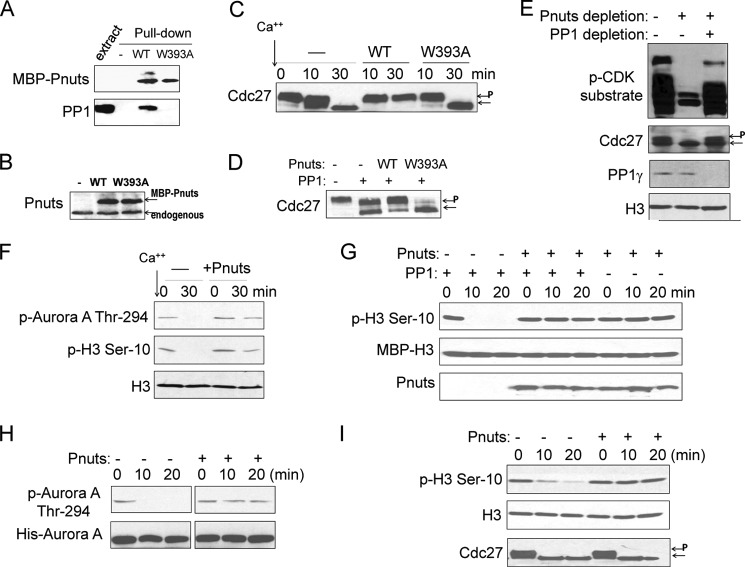
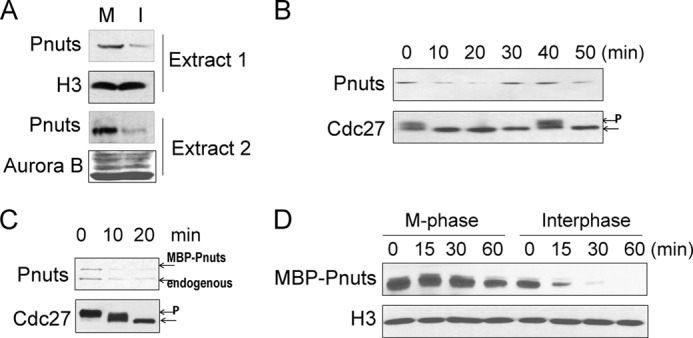
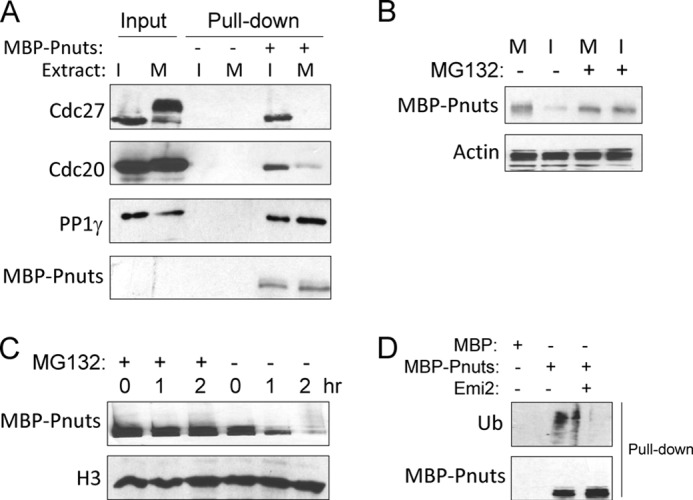
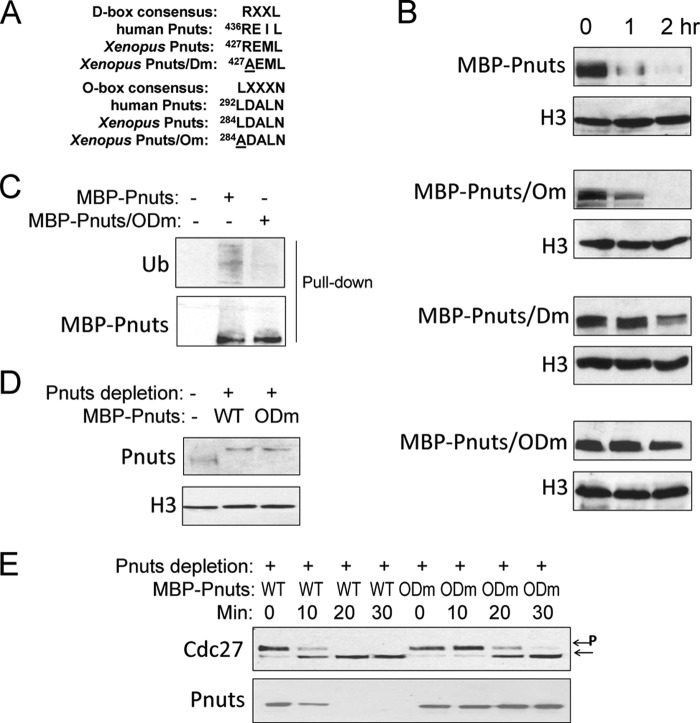
Mitotic progression is regulated largely through dynamic and reversible protein phosphorylation that is modulated by opposing actions of protein kinases and phosphatases. In this study, we show that phosphatase 1 nuclear targeting subunit (Pnuts) functions as a master regulator of mitosis by modulating protein phosphatase 1 (PP1). Overexpression of Pnuts in Xenopus egg extracts inhibited both mitotic and meiotic exit. Immunodepletion of Pnuts from egg extracts revealed its essential functions in mitotic entry and maintenance. The level of Pnuts oscillates during the cell cycle and peaks in mitosis. Pnuts destruction during M-phase exit is mediated by the anaphase-promoting complex/cyclosome (APC/C)-targeted ubiquitination and proteolysis, and conserved destruction motifs of Pnuts. Disruption of Pnuts degradation delayed M-phase exit, suggesting it as an important mechanism to permit M-phase exit.
Keywords: APC/C; Cell Cycle; Mitosis; Phosphatase; Phosphoprotein Phosphatase 1 (PP1); Pnuts; Xenopus.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Janssen A., Medema R. H. (2011) Mitosis as an anti-cancer target. Oncogene 30, 2799–2809 - PubMed
-
- Nigg E. A. (2001) Mitotic kinases as regulators of cell division and its checkpoints. Nat. Rev. Mol. Cell Biol. 2, 21–32 - PubMed
-
- Lens S. M., Voest E. E., Medema R. H. (2010) Shared and separate functions of Polo-like kinases and Aurora kinases in cancer. Nat. Rev. Cancer 10, 825–841 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
