

Multimodal Approaches for Regenerative Stroke Therapies: Combination of Granulocyte Colony-Stimulating Factor with Bone Marrow Mesenchymal Stem Cells is Not Superior to G-CSF Alone
- PMID: 25002846
- PMCID: PMC4066299
- DOI: 10.3389/fnagi.2014.00130
Multimodal Approaches for Regenerative Stroke Therapies: Combination of Granulocyte Colony-Stimulating Factor with Bone Marrow Mesenchymal Stem Cells is Not Superior to G-CSF Alone
Abstract
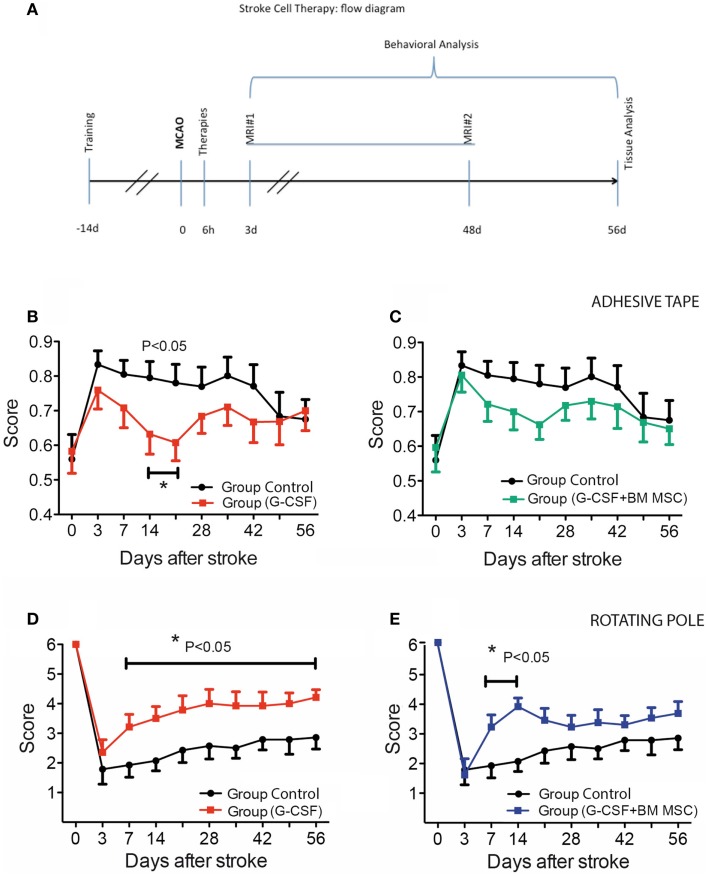
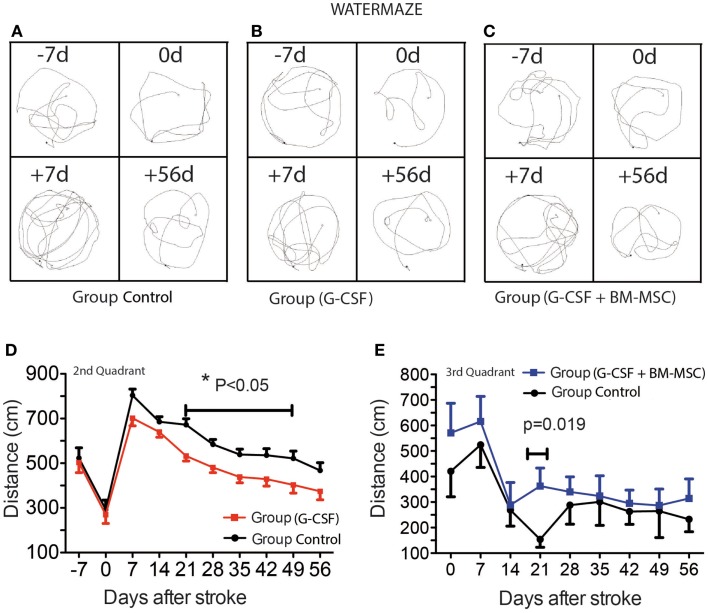
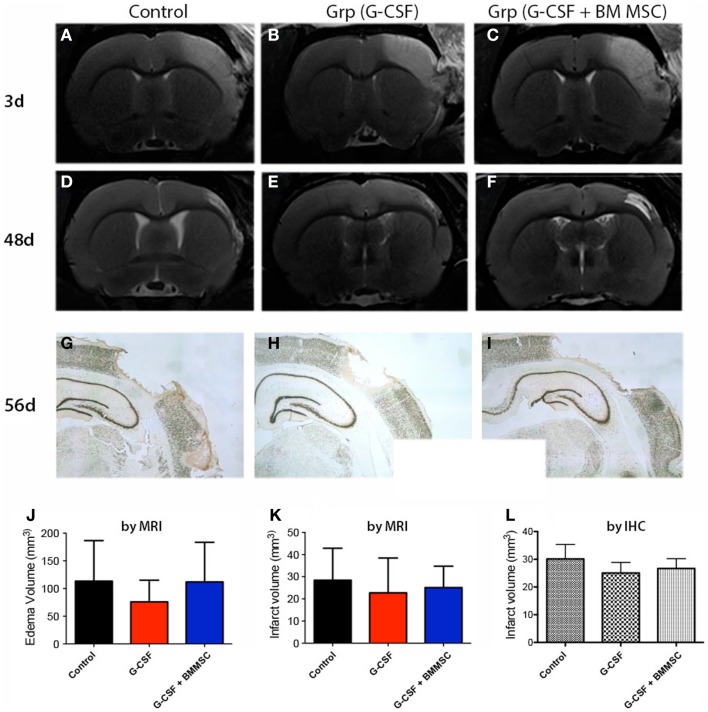
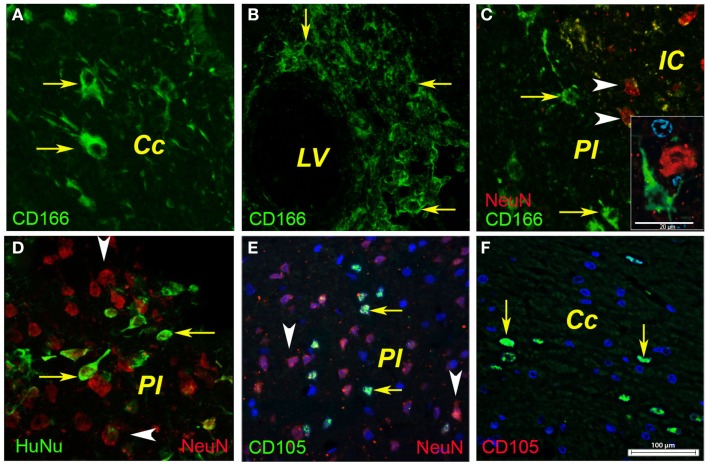
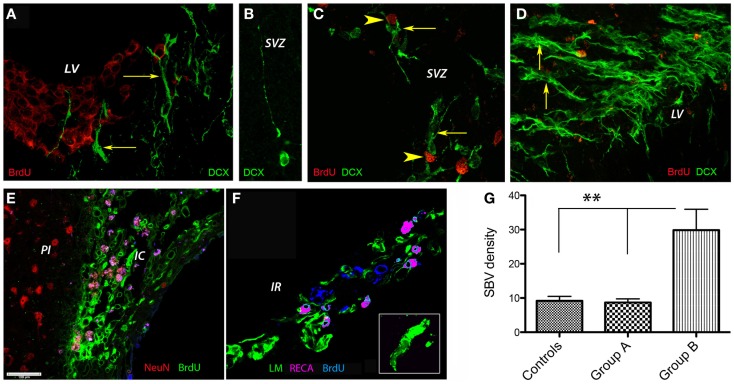
Attractive therapeutic strategies to enhance post-stroke recovery of aged brains include methods of cellular therapy that can enhance the endogenous restorative mechanisms of the injured brain. Since stroke afflicts mostly the elderly, it is highly desirable to test the efficacy of cell therapy in the microenvironment of aged brains that is generally refractory to regeneration. In particular, stem cells from the bone marrow allow an autologous transplantation approach that can be translated in the near future to the clinical practice. Such a bone marrow-derived therapy includes the grafting of stem cells as well as the delayed induction of endogenous stem cell mobilization and homing by the stem cell mobilizer granulocyte colony-stimulating factor (G-CSF). We tested the hypothesis that grafting of bone marrow-derived pre-differentiated mesenchymal cells (BM-MSCs) in G-CSF-treated animals improves the long-term functional outcome in aged rodents. To this end, G-CSF alone (50 μg/kg) or in combination with a single dose (10(6) cells) of rat BM MSCs was administered intravenously to Sprague-Dawley rats at 6 h after transient occlusion (90 min) of the middle cerebral artery. Infarct volume was measured by magnetic resonance imaging at 3 and 48 days post-stroke and additionally by immunhistochemistry at day 56. Functional recovery was tested during the entire post-stroke survival period of 56 days. Daily treatment for post-stroke aged rats with G-CSF led to a robust and consistent improvement of neurological function after 28 days. The combination therapy also led to robust angiogenesis in the formerly infarct core and beyond in the "islet of regeneration." However, G-CSF + BM MSCs may not impact at all on the spatial reference-memory task or infarct volume and therefore did not further improve the post-stroke recovery. We suggest that in a real clinical practice involving older post-stroke patients, successful regenerative therapies would have to be carried out for a much longer time.
Keywords: BM MSC; G-CSF; aging; angiogenesis; cell therapy; stroke; translational medicine.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources