

Budding yeast kinetochore proteins, Chl4 and Ctf19, are required to maintain SPB-centromere proximity during G1 and late anaphase
- PMID: 25003500
- PMCID: PMC4086815
- DOI: 10.1371/journal.pone.0101294
Budding yeast kinetochore proteins, Chl4 and Ctf19, are required to maintain SPB-centromere proximity during G1 and late anaphase
Abstract
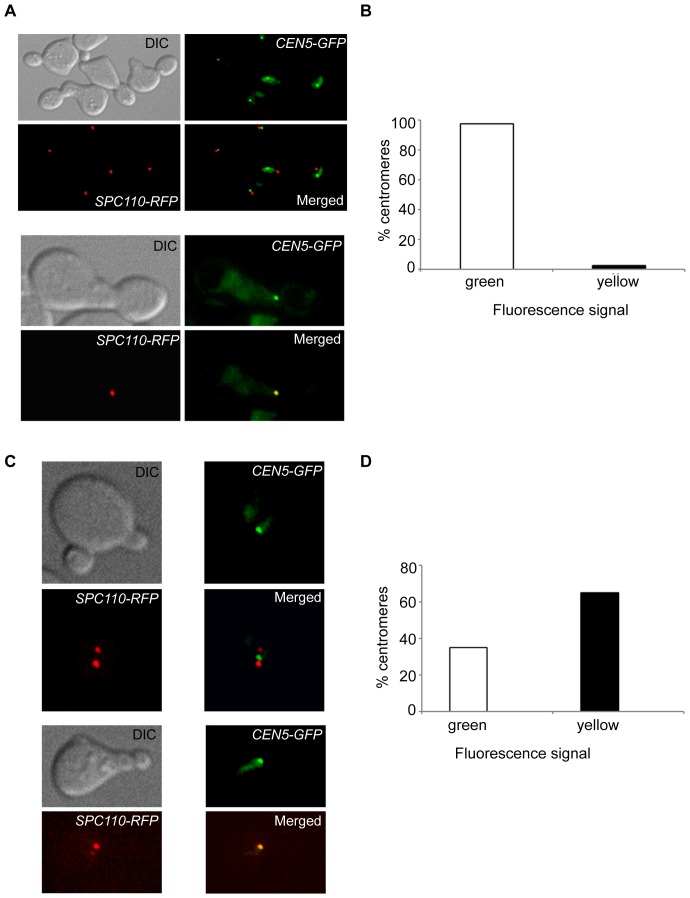
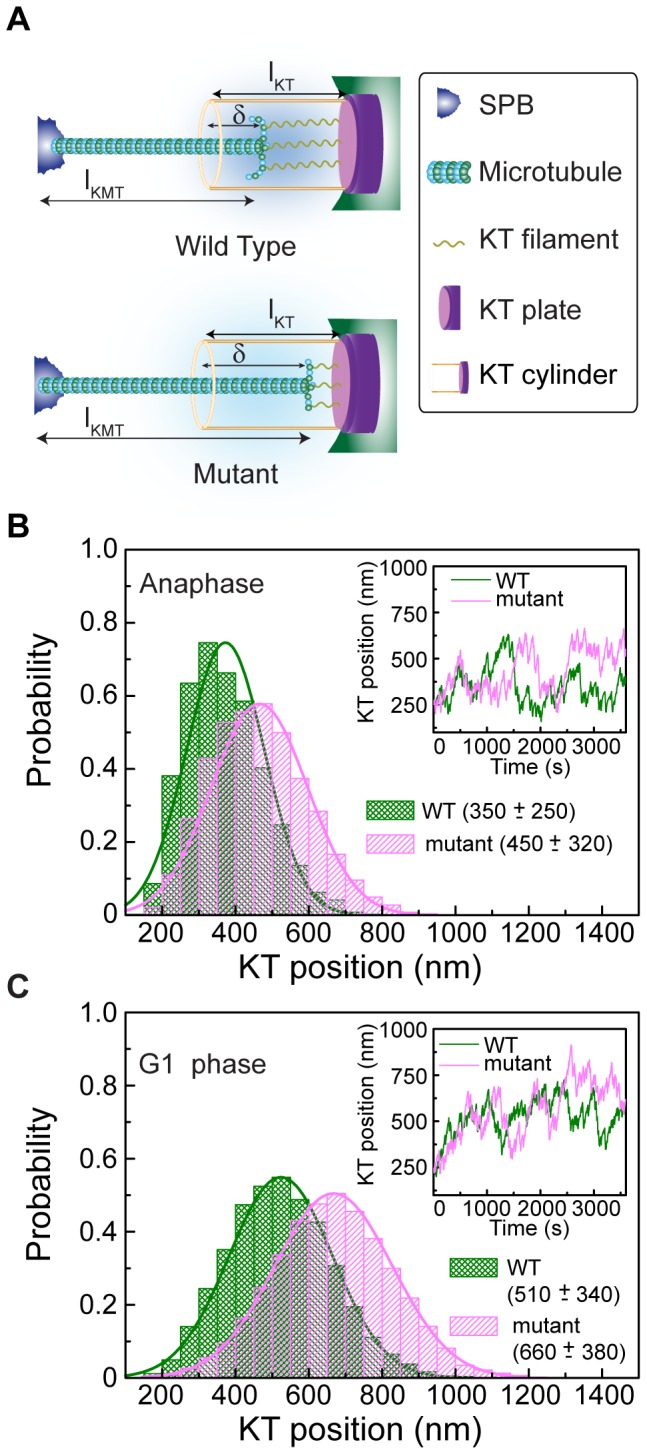
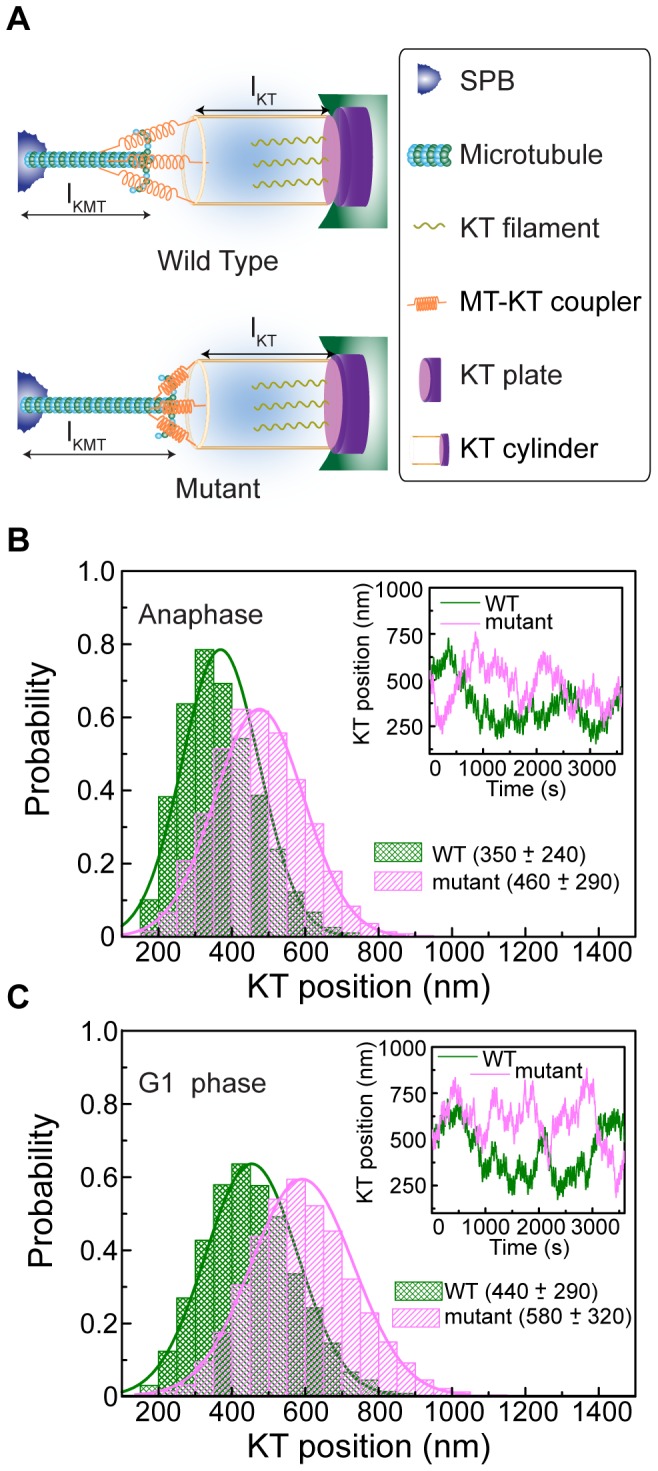
In the budding yeast, centromeres stay clustered near the spindle pole bodies (SPBs) through most of the cell cycle. This SPB-centromere proximity requires microtubules and functional kinetochores, which are protein complexes formed on the centromeres and capable of binding microtubules. The clustering is suggested by earlier studies to depend also on protein-protein interactions between SPB and kinetochore components. Previously it has been shown that the absence of non-essential kinetochore proteins of the Ctf19 complex weakens kinetochore-microtubule interaction, but whether this compromised interaction affects centromere/kinetochore positioning inside the nucleus is unknown. We found that in G1 and in late anaphase, SPB-centromere proximity was disturbed in mutant cells lacking Ctf19 complex members,Chl4p and/or Ctf19p, whose centromeres lay further away from their SPBs than those of the wild-type cells. We unequivocally show that the SPB-centromere proximity and distances are not dependent on physical interactions between SPB and kinetochore components, but involve microtubule-dependent forces only. Further insight on the positional difference between wild-type and mutant kinetochores was gained by generating computational models governed by (1) independently regulated, but constant kinetochore microtubule (kMT) dynamics, (2) poleward tension on kinetochore and the antagonistic polar ejection force and (3) length and force dependent kMT dynamics. Numerical data obtained from the third model concurs with experimental results and suggests that the absence of Chl4p and/or Ctf19p increases the penetration depth of a growing kMT inside the kinetochore and increases the rescue frequency of a depolymerizing kMT. Both the processes result in increased distance between SPB and centromere.
Conflict of interest statement
Figures


Similar articles
-
Iml3p, a component of the Ctf19 complex of the budding yeast kinetochore is required to maintain kinetochore integrity under conditions of spindle stress.FEMS Yeast Res. 2013 Jun;13(4):375-85. doi: 10.1111/1567-1364.12041. Epub 2013 Apr 2. FEMS Yeast Res. 2013. PMID: 23480593
-
Chl4p and iml3p are two new members of the budding yeast outer kinetochore.Mol Biol Cell. 2003 Feb;14(2):460-76. doi: 10.1091/mbc.e02-08-0517. Mol Biol Cell. 2003. PMID: 12589047 Free PMC article.
-
Four new subunits of the Dam1-Duo1 complex reveal novel functions in sister kinetochore biorientation.EMBO J. 2002 Jan 15;21(1-2):181-93. doi: 10.1093/emboj/21.1.181. EMBO J. 2002. PMID: 11782438 Free PMC article.
-
Structures and functions of yeast kinetochore complexes.Annu Rev Biochem. 2007;76:563-91. doi: 10.1146/annurev.biochem.76.052705.160607. Annu Rev Biochem. 2007. PMID: 17362199 Review.
-
Motile kinetochores and polar ejection forces dictate chromosome position on the vertebrate mitotic spindle.J Cell Biol. 1994 Feb;124(3):223-33. doi: 10.1083/jcb.124.3.223. J Cell Biol. 1994. PMID: 8294508 Free PMC article. Review.
Cited by
-
A comprehensive model to predict mitotic division in budding yeasts.Mol Biol Cell. 2015 Nov 5;26(22):3954-65. doi: 10.1091/mbc.E15-04-0236. Epub 2015 Aug 26. Mol Biol Cell. 2015. PMID: 26310442 Free PMC article.
-
Spatio-temporal regulation of nuclear division by Aurora B kinase Ipl1 in Cryptococcus neoformans.PLoS Genet. 2019 Feb 14;15(2):e1007959. doi: 10.1371/journal.pgen.1007959. eCollection 2019 Feb. PLoS Genet. 2019. PMID: 30763303 Free PMC article.
-
Evidence of 14-3-3 proteins contributing to kinetochore integrity and chromosome congression during mitosis.J Cell Sci. 2024 Aug 1;137(15):jcs261928. doi: 10.1242/jcs.261928. Epub 2024 Aug 9. J Cell Sci. 2024. PMID: 38988319 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
