

Formation of cartilage and synovial tissue by human gingival stem cells
- PMID: 25003637
- PMCID: PMC4236036
- DOI: 10.1089/scd.2013.0547
Formation of cartilage and synovial tissue by human gingival stem cells
Abstract
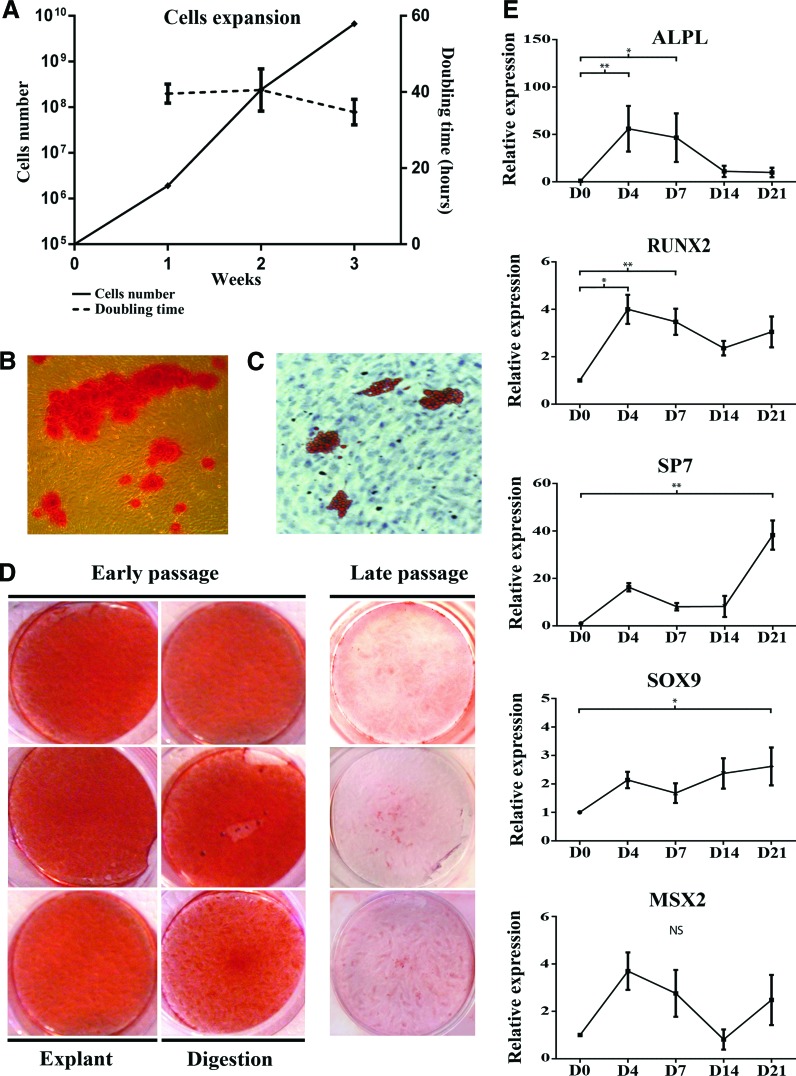
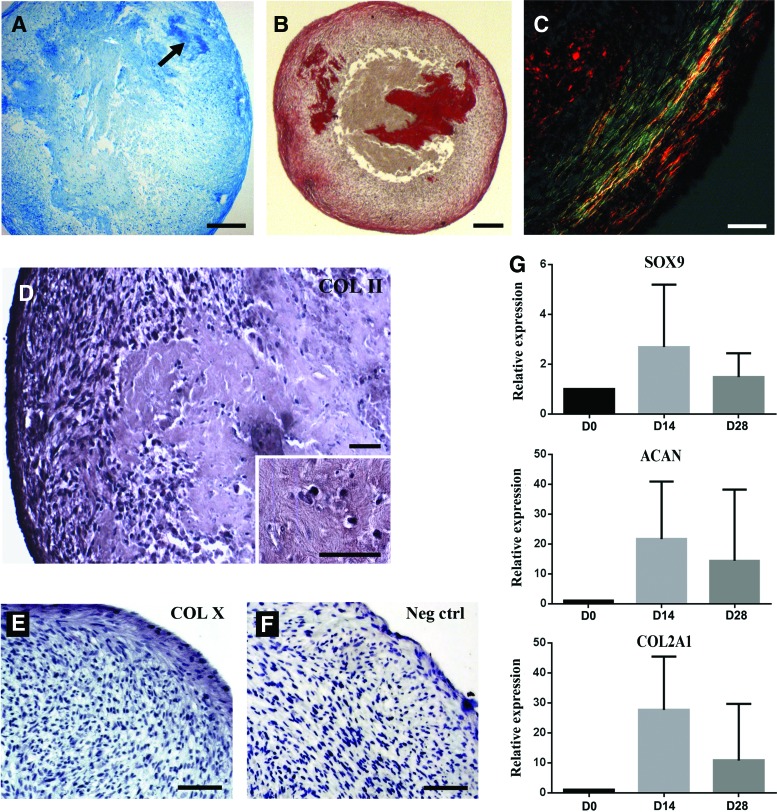
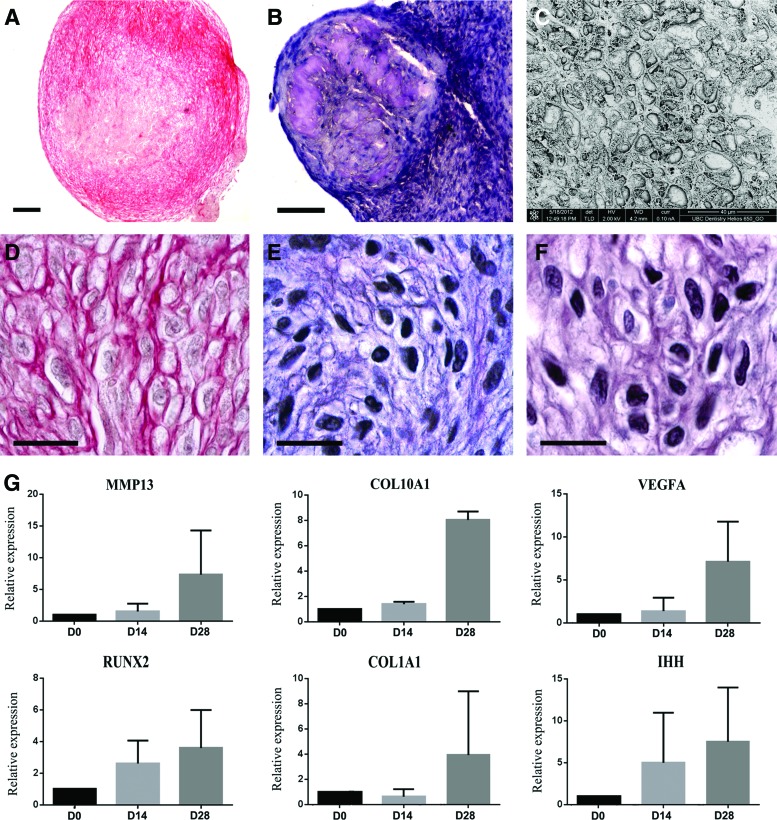
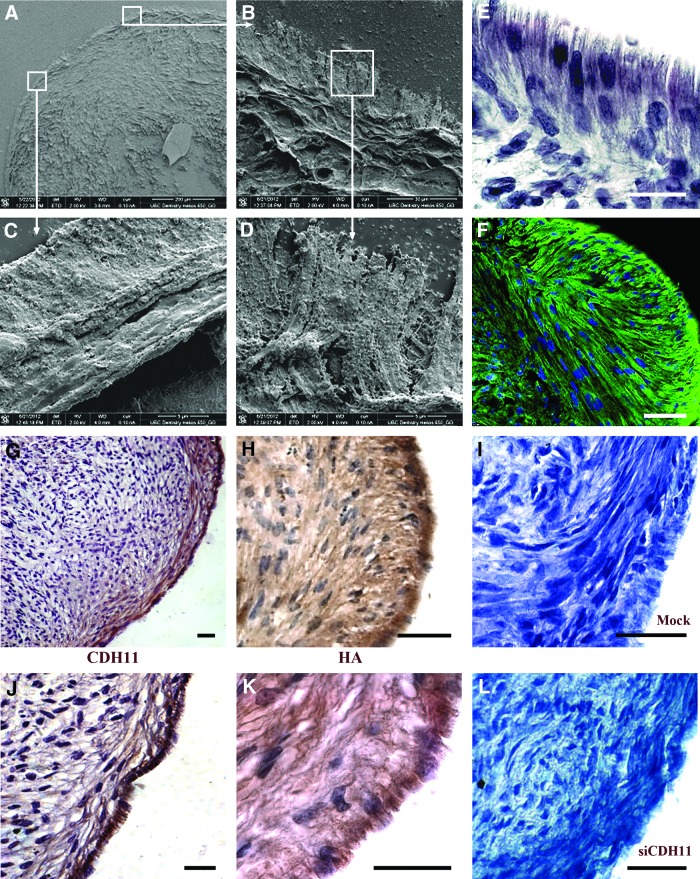
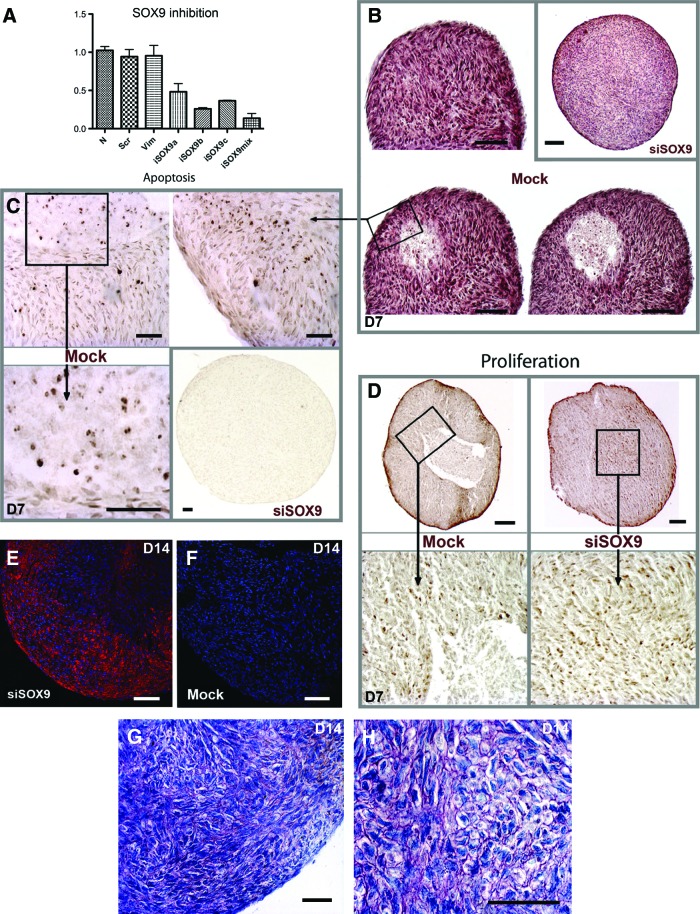
Human gingival stem cells (HGSCs) can be easily isolated and manipulated in culture to investigate their multipotency. Osteogenic differentiation of bone-marrow-derived mesenchymal stem/stromal cells has been well documented. HGSCs derive from neural crests, however, and their differentiation capacity has not been fully established. The aim of the present report was to investigate whether HGSCs can be induced to differentiate to osteoblasts and chondrocytes. HGSCs were cultured either in a classical monolayer culture or in three-dimensional floating micromass pellet cultures in specific differentiation media. HGSC differentiation to osteogenic and chondrogenic lineages was determined by protein and gene expression analyses, and also by specific staining of cells and tissue pellets. HGSCs cultured in osteogenic differentiation medium showed induction of Runx2, alkaline phosphatase (ALPL), and osterix expression, and subsequently formed mineralized nodules consistent with osteogenic differentiation. Interestingly, HGSC micromass cultures maintained in chondrogenic differentiation medium showed SOX9-dependent differentiation to both chondrocyte and synoviocyte lineages. Chondrocytes at different stages of differentiation were identified by gene expression profiles and by histochemical and immunohistochemical staining. In 3-week-old cultures, peripheral cells in the micromass cultures organized in layers of cuboidal cells with villous structures facing the medium. These cells were strongly positive for cadherin-11, a marker of synoviocytes. In summary, the findings indicate that HGSCs have the capacity to differentiate to osteogenic, chondrogenic, and synoviocyte lineages. Therefore, HGSCs could serve as an alternative source for stem cell therapies in regenerative medicine for patients with cartilage and joint destructions, such as observed in rheumatoid arthritis.
Figures
Similar articles
-
Perlecan is required for the chondrogenic differentiation of synovial mesenchymal cells through regulation of Sox9 gene expression.J Orthop Res. 2017 Apr;35(4):837-846. doi: 10.1002/jor.23318. Epub 2016 Jun 15. J Orthop Res. 2017. PMID: 27238423
-
In-vitro chondrogenic potential of synovial stem cells and chondrocytes allocated for autologous chondrocyte implantation - a comparison : Synovial stem cells as an alternative cell source for autologous chondrocyte implantation.Int Orthop. 2017 May;41(5):991-998. doi: 10.1007/s00264-017-3400-y. Epub 2017 Jan 25. Int Orthop. 2017. PMID: 28124102
-
Mesenchymal stem cell-derived extracellular matrix enhances chondrogenic phenotype of and cartilage formation by encapsulated chondrocytes in vitro and in vivo.Acta Biomater. 2018 Mar 15;69:71-82. doi: 10.1016/j.actbio.2017.12.043. Epub 2018 Jan 6. Acta Biomater. 2018. PMID: 29317369 Free PMC article.
-
Generation of human induced pluripotent stem cells from osteoarthritis patient-derived synovial cells.Arthritis Rheum. 2011 Oct;63(10):3010-21. doi: 10.1002/art.30488. Arthritis Rheum. 2011. PMID: 21953087
-
Tissue engineering of the synovial joint: the role of cell density.Proc Inst Mech Eng H. 2007 Jul;221(5):429-40. doi: 10.1243/09544119JEIM288. Proc Inst Mech Eng H. 2007. PMID: 17822145 Review.
Cited by
-
Gingival epithelial cell-derived microvesicles activate mineralization in gingival fibroblasts.Sci Rep. 2022 Sep 22;12(1):15779. doi: 10.1038/s41598-022-19732-1. Sci Rep. 2022. PMID: 36138045 Free PMC article.
-
[Research advances on the mechanism of oral mucosal stem cells in promoting wound healing].Zhonghua Shao Shang Yu Chuang Mian Xiu Fu Za Zhi. 2023 May 20;39(5):496-500. doi: 10.3760/cma.j.cn501225-20220730-00321. Zhonghua Shao Shang Yu Chuang Mian Xiu Fu Za Zhi. 2023. PMID: 37805762 Free PMC article. Review. Chinese.
-
Contribution of neural crest-derived stem cells and nasal chondrocytes to articular cartilage regeneration.Cell Mol Life Sci. 2020 Dec;77(23):4847-4859. doi: 10.1007/s00018-020-03567-y. Epub 2020 Jun 5. Cell Mol Life Sci. 2020. PMID: 32504256 Free PMC article. Review.
-
Effects of High-Temperature-Pressure Polymerized Resin-Infiltrated Ceramic Networks on Oral Stem Cells.PLoS One. 2016 May 19;11(5):e0155450. doi: 10.1371/journal.pone.0155450. eCollection 2016. PLoS One. 2016. PMID: 27196425 Free PMC article.
-
3D Bioprinting of Hyaline Cartilage Using Nasal Chondrocytes.Ann Biomed Eng. 2024 Jul;52(7):1816-1834. doi: 10.1007/s10439-023-03176-3. Epub 2023 Mar 23. Ann Biomed Eng. 2024. PMID: 36952145 Review.
References
-
- Scotti C, Tonnarelli B, Papadimitropoulos A, Scherberich A, Schaeren S, Schauerte A, Lopez-Rios J, Zeller R, Barbero A. and Martin I. (2010). Recapitulation of endochondral bone formation using human adult mesenchymal stem cells as a paradigm for developmental engineering. Proc Natl Acad Sci U S A 107:7251–7256 - PMC - PubMed
-
- Ornitz DM. and Marie PJ. (2002). FGF signaling pathways in endochondral and intramembranous bone development and human genetic disease. Genes Dev 16:1446–1465 - PubMed
-
- Kronenberg HM. (2003). Developmental regulation of the growth plate. Nature 423:332–336 - PubMed
-
- Archer CW, Dowthwaite GP. and Francis-West P. (2003). Development of synovial joints. Birth Defects Res C Embryo Today 69:144–155 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
