

Resolution of acute inflammation bridges the gap between innate and adaptive immunity
- PMID: 25006125
- PMCID: PMC4383794
- DOI: 10.1182/blood-2014-03-562710
Resolution of acute inflammation bridges the gap between innate and adaptive immunity
Abstract
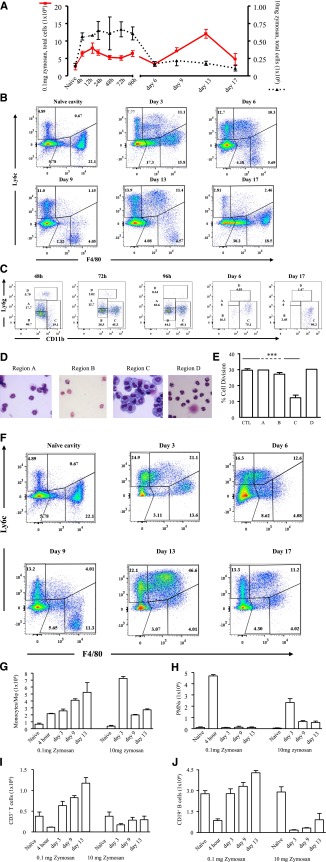
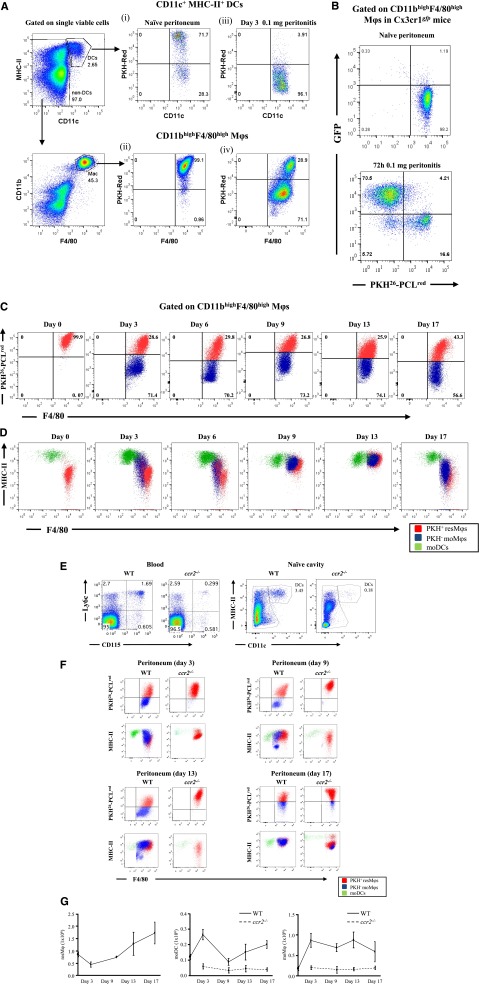
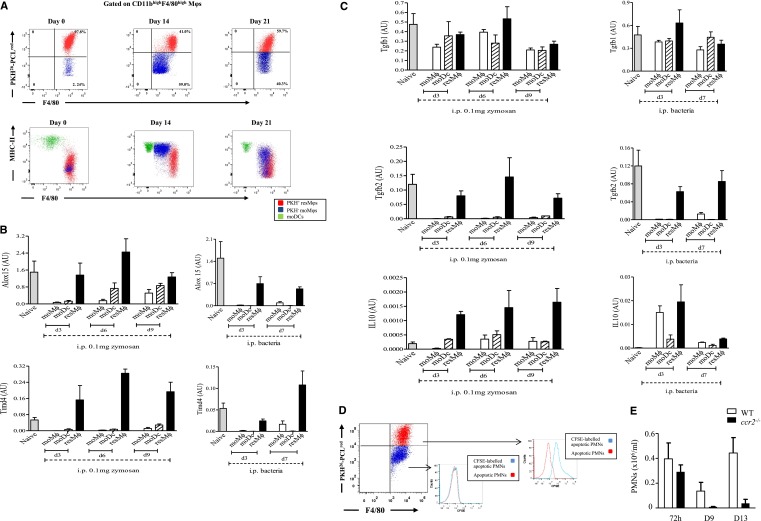
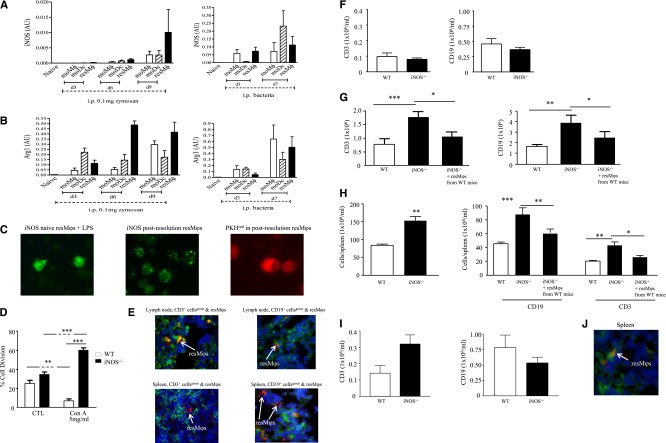
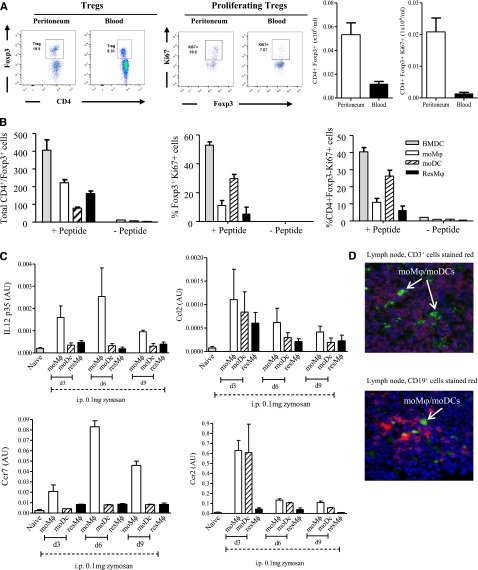
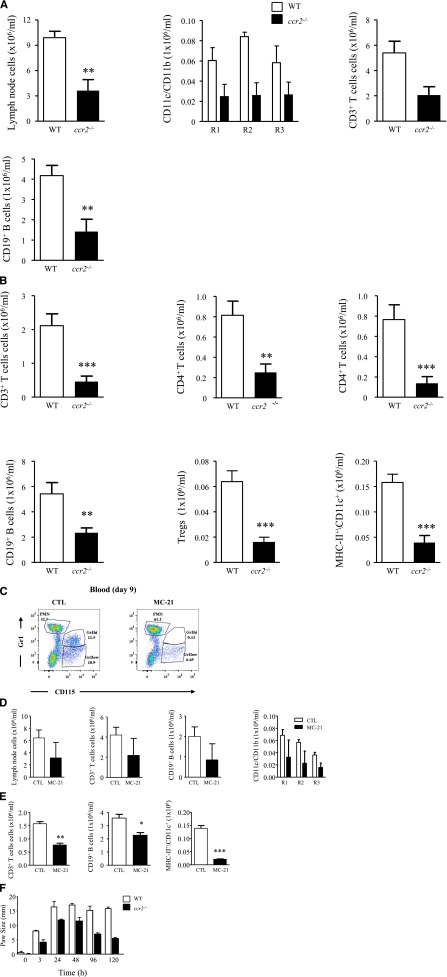
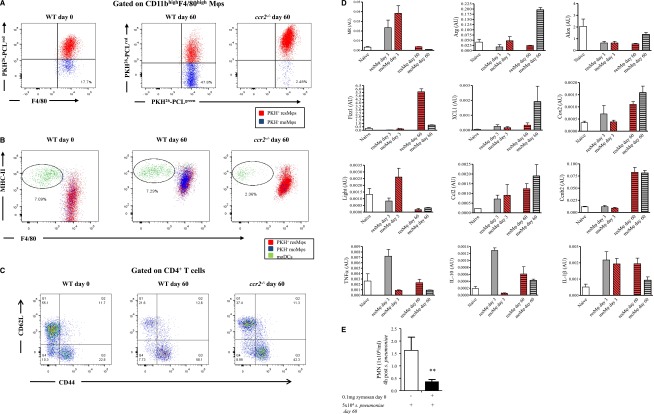
Acute inflammation is traditionally characterized by polymorphonuclear leukocytes (PMN) influx followed by phagocytosing macrophage (Mφs) that clear injurious stimuli leading to resolution and tissue homeostasis. However, using the peritoneal cavity, we found that although innate immune-mediated responses to low-dose zymosan or bacteria resolve within days, these stimuli, but not hyperinflammatory stimuli, trigger a previously overlooked second wave of leukocyte influx into tissues that persists for weeks. These cells comprise distinct populations of tissue-resident Mφs (resMφs), Ly6c(hi) monocyte-derived Mφs (moMφs), monocyte-derived dendritic cells (moDCs), and myeloid-derived suppressor cells (MDSCs). Postresolution mononuclear phagocytes were observed alongside lymph node expansion and increased numbers of blood and peritoneal memory T and B lymphocytes. The resMφs and moMφs triggered FoxP3 expression within CD4 cells, whereas moDCs drive T-cell proliferation. The resMφs preferentially clear apoptotic PMNs and migrate to lymph nodes to bring about their contraction in an inducible nitric oxide synthase-dependent manner. Finally, moMφs remain in tissues for months postresolution, alongside altered numbers of T cells collectively dictating the magnitude of subsequent acute inflammatory reactions. These data challenge the prevailing idea that resolution leads back to homeostasis and asserts that resolution acts as a bridge between innate and adaptive immunity, as well as tissue reprogramming.
© 2014 by The American Society of Hematology.
Figures
Comment in
-
Cellular dynamics of resolving inflammation.Blood. 2014 Sep 11;124(11):1701-3. doi: 10.1182/blood-2014-07-589341. Blood. 2014. PMID: 25214196 Free PMC article.
References
-
- Serhan CN, Savill J. Resolution of inflammation: the beginning programs the end. Nat Immunol. 2005;6(12):1191–1197. - PubMed
-
- Jamieson T, Cook DN, Nibbs RJ, et al. The chemokine receptor D6 limits the inflammatory response in vivo. Nat Immunol. 2005;6(4):403–411. - PubMed
-
- Lohr J, Knoechel B, Abbas AK. Regulatory T cells in the periphery. Immunol Rev. 2006;212:149–162. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
