

G1-arrested newborn cells are the predominant infectious form of the pathogen Brucella abortus
- PMID: 25006695
- PMCID: PMC4104442
- DOI: 10.1038/ncomms5366
G1-arrested newborn cells are the predominant infectious form of the pathogen Brucella abortus
Abstract
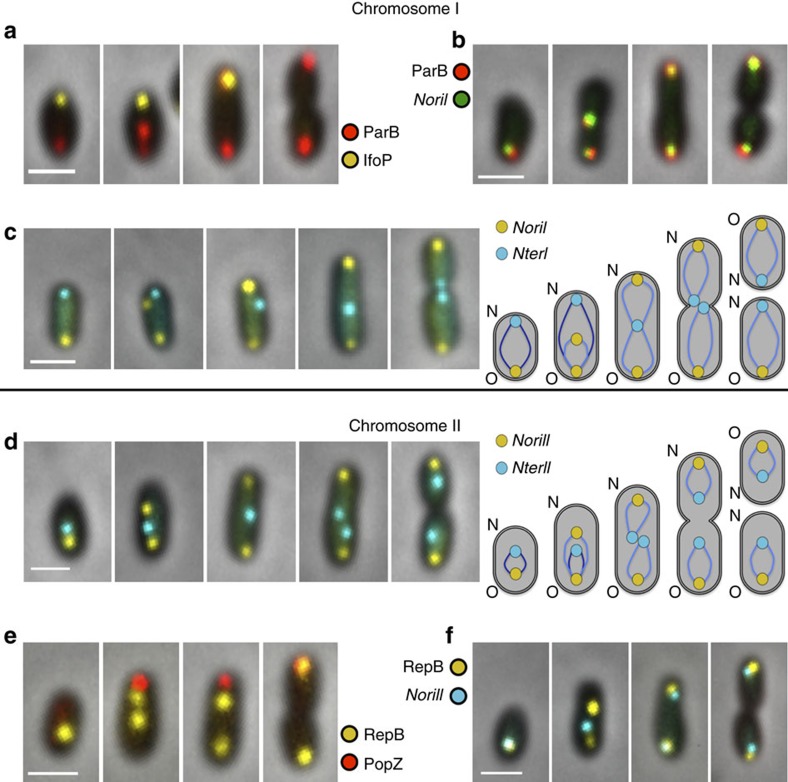
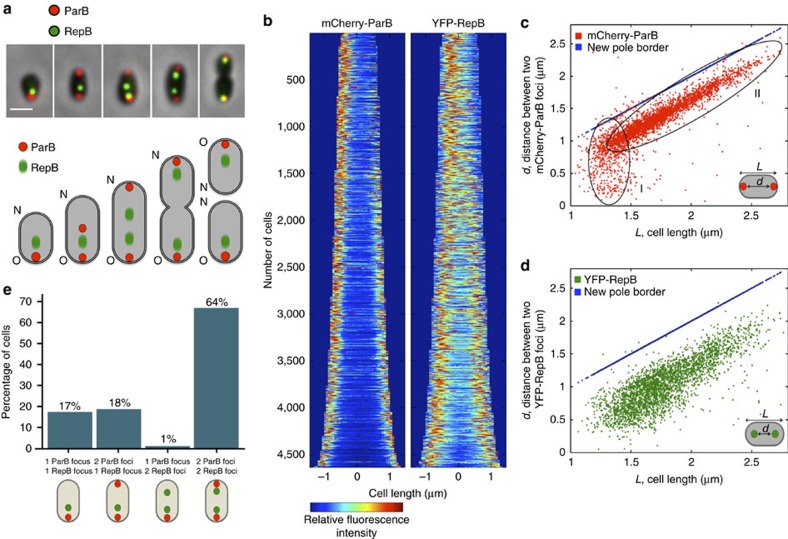
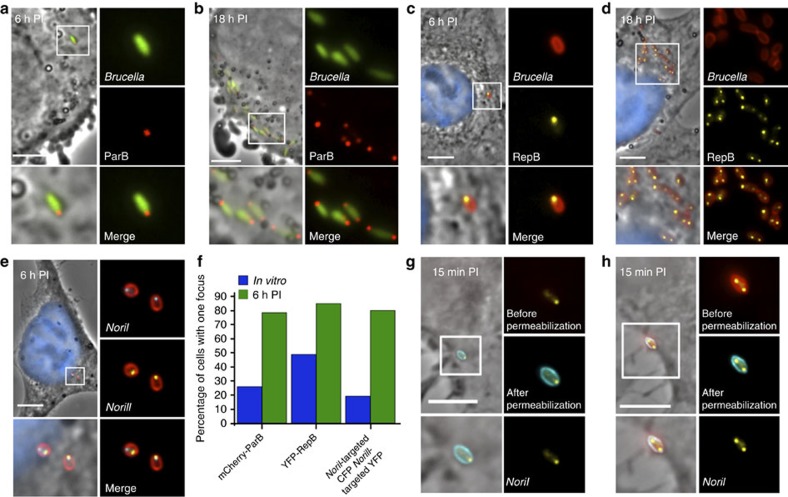
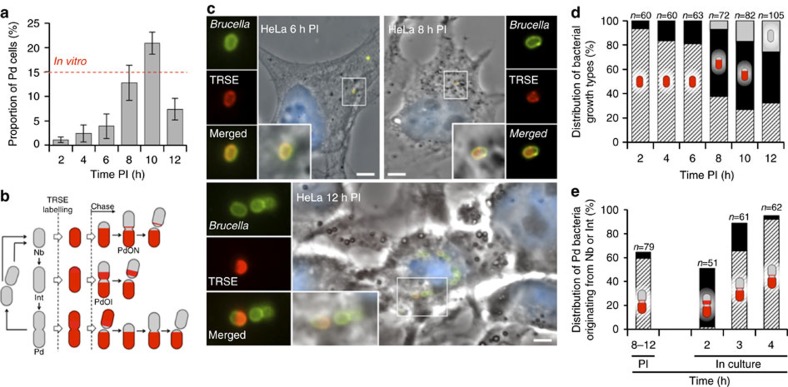
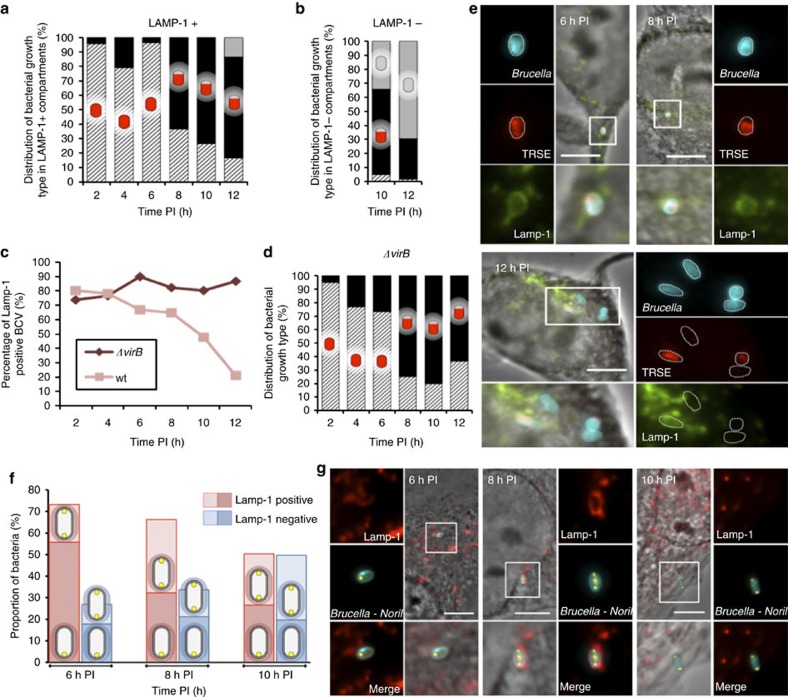
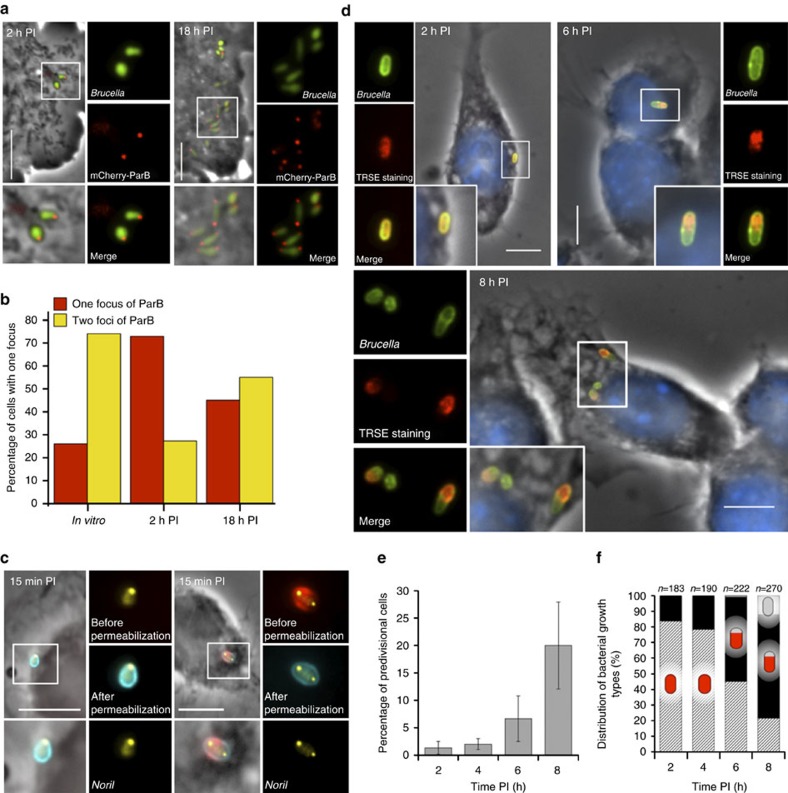
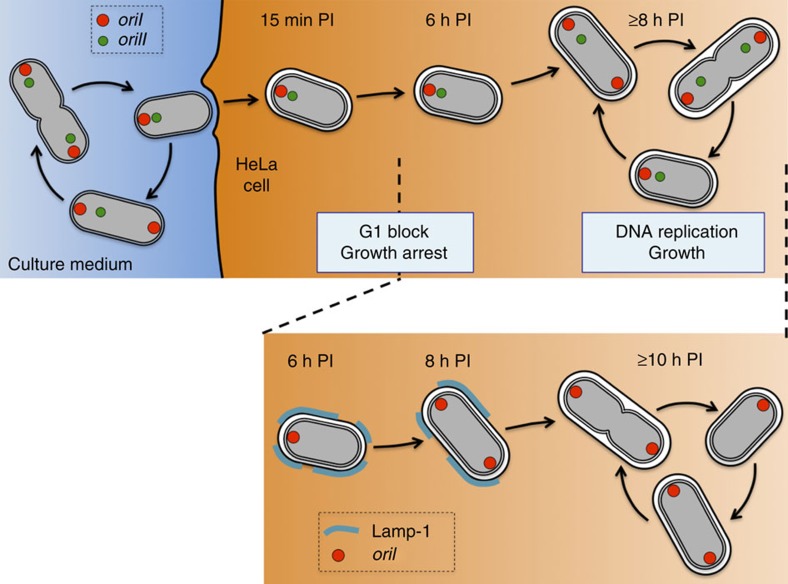
Several intracellular pathogens, such as Brucella abortus, display a biphasic infection process starting with a non-proliferative stage of unclear nature. Here, we study the cell cycle of B. abortus at the single-cell level, in culture and during infection of HeLa cells and macrophages. The localization of segregation and replication loci of the two bacterial chromosomes indicates that, immediately after being engulfed by host-cell endocytic vacuoles, most bacterial cells are newborn. These bacterial cells do not initiate DNA replication for the next 4 to 6 h, indicating a G1 arrest. Moreover, growth is completely stopped during that time, reflecting a global cell cycle block. Growth and DNA replication resume later, although bacteria still reside within endosomal-like compartments. We hypothesize that the predominance of G1-arrested bacteria in the infectious population, and the bacterial cell cycle arrest following internalization, may constitute a widespread strategy among intracellular pathogens to colonize new proliferation niches.
Figures
Comment in
-
On the link between cell cycle and infection of the Alphaproteobacterium Brucella abortus.Microb Cell. 2014 Sep 29;1(10):346-348. doi: 10.15698/mic2014.10.171. Microb Cell. 2014. PMID: 28357212 Free PMC article.
References
-
- Salcedo S. P. & Holden D. W. Bacterial interactions with the eukaryotic secretory pathway. Curr. Opin. Microbiol. 8, 92–98 (2005). - PubMed
-
- Moreno E. & Moriyon I. Prokaryotes 5, 315–456Springer (2006).
-
- Comerci D. J., Martinez-Lorenzo M. J., Sieira R., Gorvel J. P. & Ugalde R. A. Essential role of the VirB machinery in the maturation of the Brucella abortus-containing vacuole. Cell. Microbiol. 3, 159–168 (2001). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
