

Impact of sequence variation in a dominant HLA-A*02-restricted epitope in hepatitis C virus on priming and cross-reactivity of CD8+ T cells
- PMID: 25008925
- PMCID: PMC4178828
- DOI: 10.1128/JVI.01590-14
Impact of sequence variation in a dominant HLA-A*02-restricted epitope in hepatitis C virus on priming and cross-reactivity of CD8+ T cells
Abstract
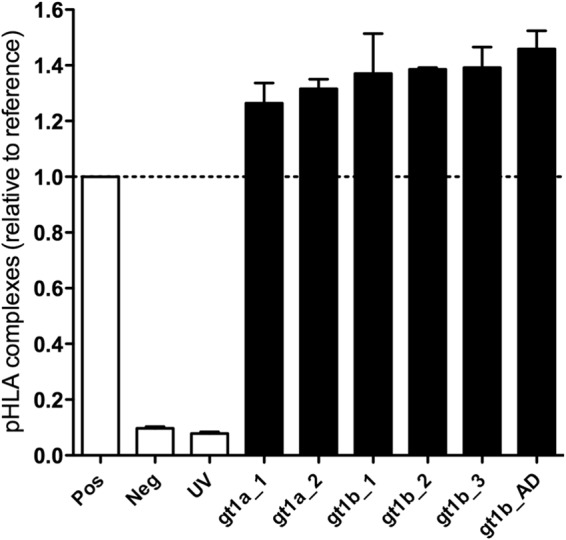
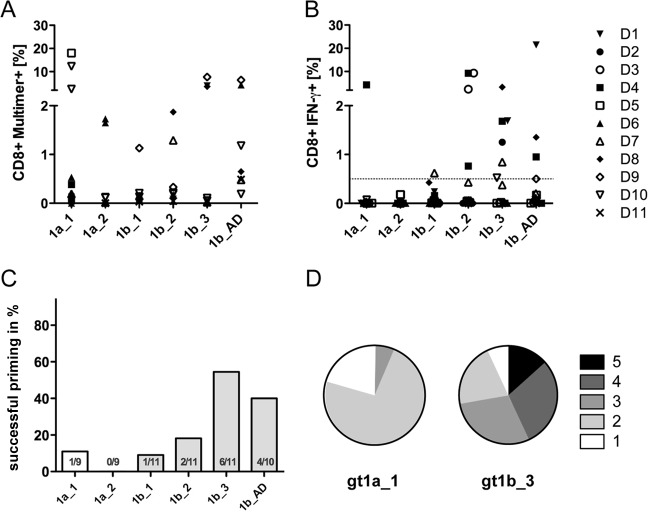
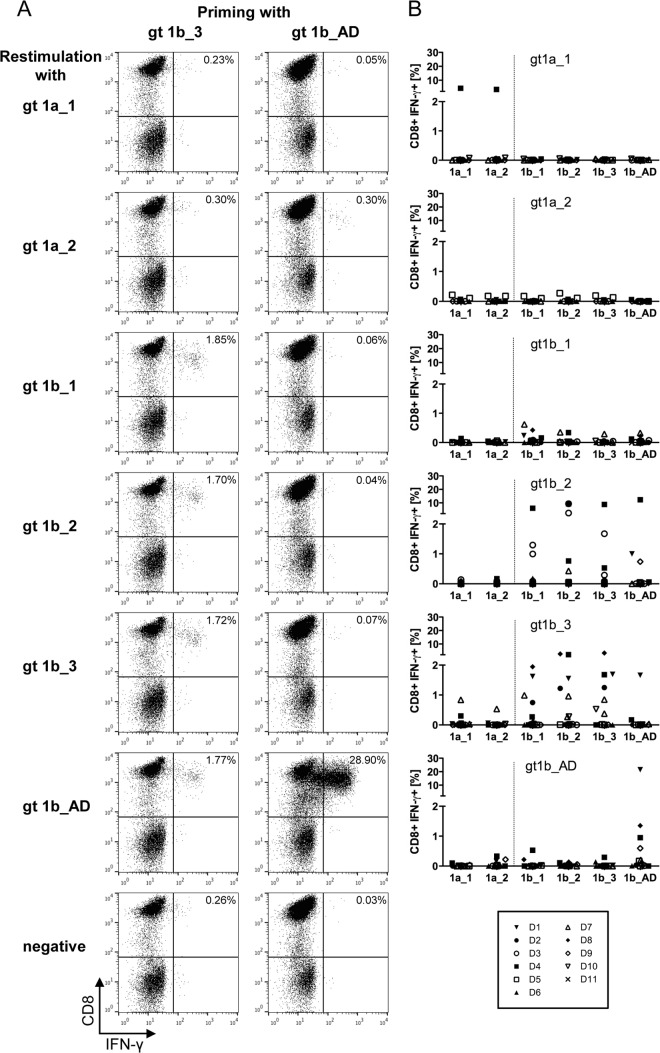
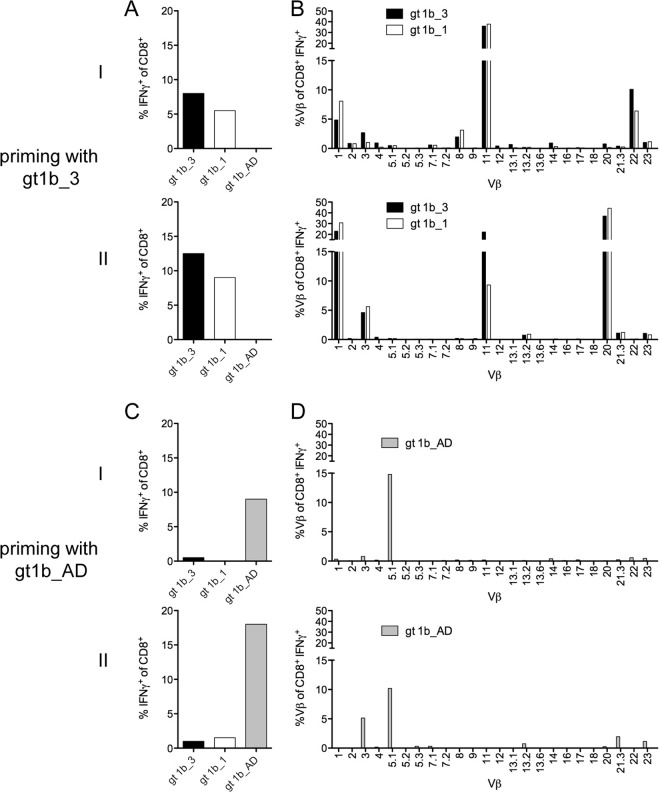
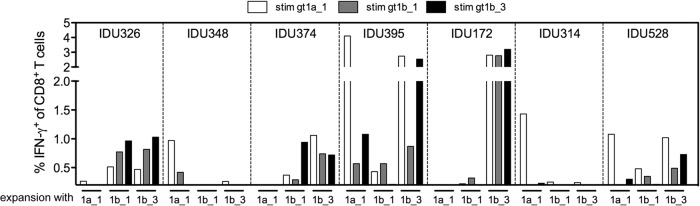
CD8+ T cells are an essential component of successful adaptive immune responses against hepatitis C virus (HCV). A major obstacle to vaccine design against HCV is its inherent viral sequence diversity. Here, we test the hypothesis that different sequence variants of an immunodominant CD8+ T cell epitope, all binding with high affinity to HLA class I, target different T cell receptor repertoires and thereby influence the quality of the CD8+ T cell response. The impacts of sequence differences in the HLA-A*02-restricted HCV NS31406-1415 epitope on in vitro priming of naive CD8+ T cells from seronegative donors and cross-reactivity of primed T cells with other epitope variants were characterized. Although the six epitope variants tested were all high-affinity binders to HLA-A*02:01, substantial differences in priming and cross-reactivity of CD8+ T cells were observed. The variant associated with the most reproducible priming and induction of T cells with broad cross-reactivity was a genotype 1b variant (KLSALGLNAV) that is more common in HCV isolates collected in Asia but is rare in sequences from Europe and North America. The superior immunogenicity and cross-reactivity of this relatively rare epitope variant were confirmed by using HCV-specific memory CD8+ T cells from people who inject drugs, who are frequently exposed to HCV. Collectively, the data suggest that sequence differences at the epitope level between HCV isolates substantially impact CD8+ T cell priming and the degree of cross-reactivity with other epitope variants.
Importance: The results have important implications for vaccine design against highly variable pathogens and suggest that evidence-based selection of the vaccine antigen sequence may improve immunogenicity and T cell cross-reactivity. Cross-reactive CD8+ T cells are likely beneficial for immune control of transmitted viruses carrying epitope variants and for prevention of immune escape during acute infection. To this end, rare epitope variants and potentially even altered epitope sequences associated with priming of broadly cross-reactive T cell receptors should be considered for vaccine design and need further testing.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
