

Structure analysis of the major capsid proteins of human polyomaviruses 6 and 7 reveals an obstructed sialic acid binding site
- PMID: 25008942
- PMCID: PMC4178836
- DOI: 10.1128/JVI.01084-14
Structure analysis of the major capsid proteins of human polyomaviruses 6 and 7 reveals an obstructed sialic acid binding site
Abstract
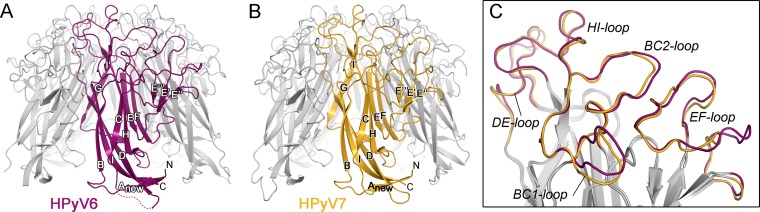
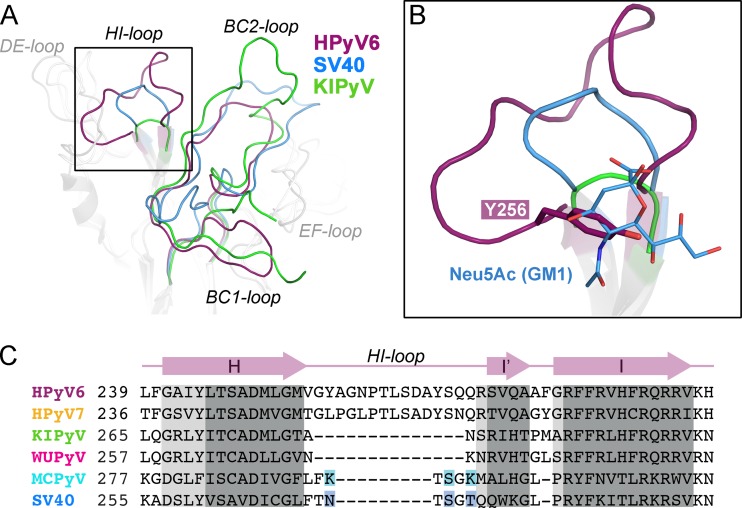
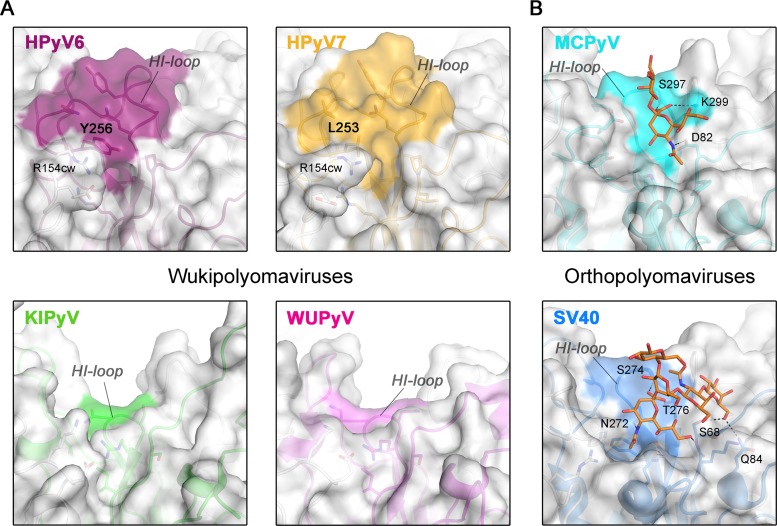
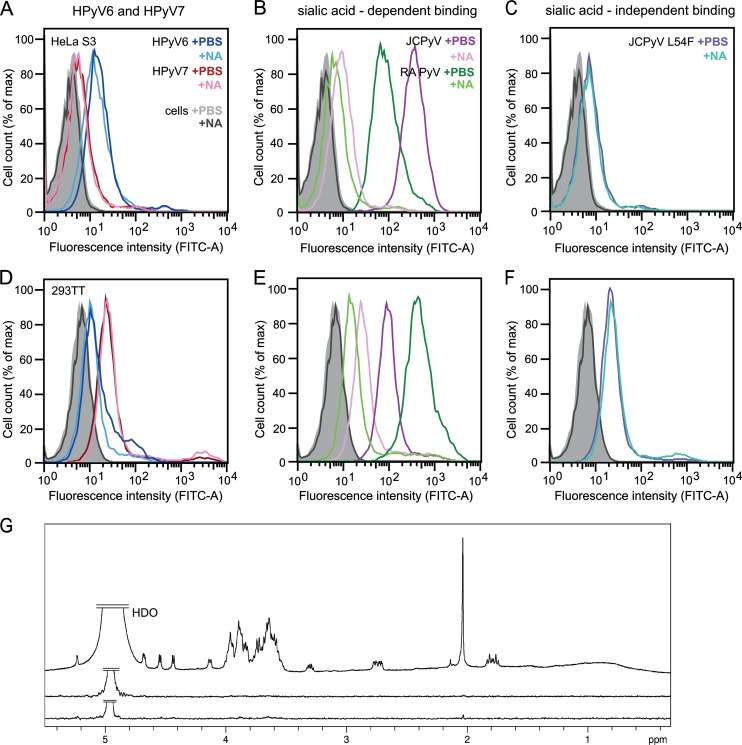
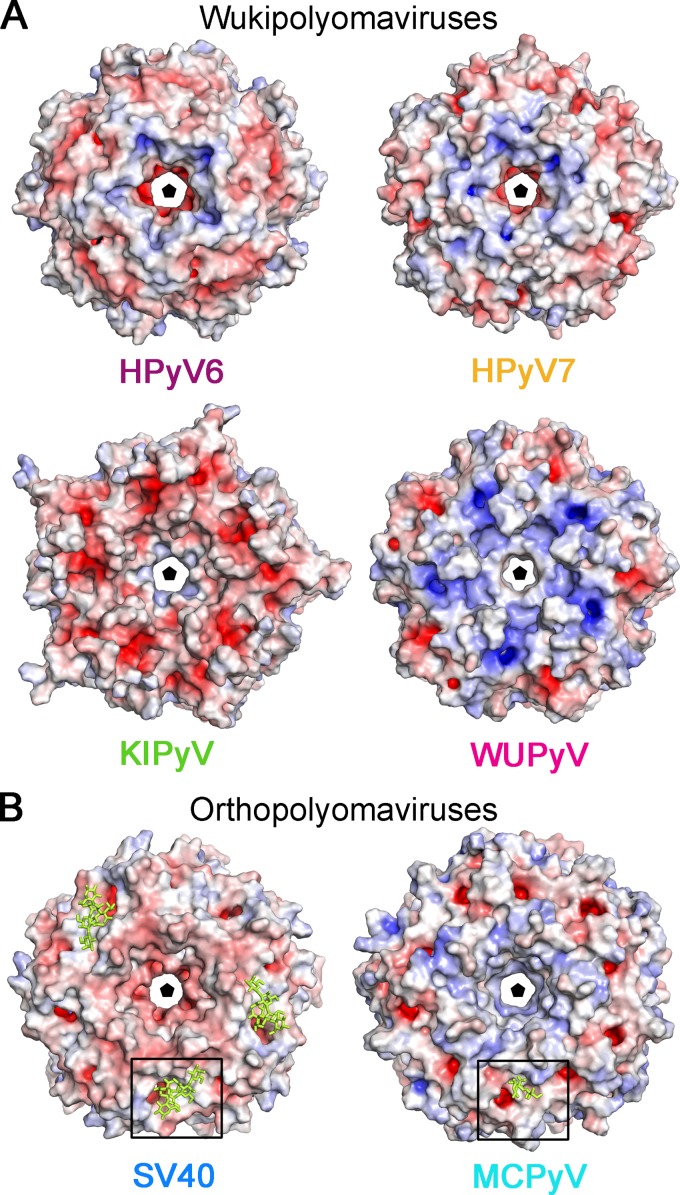
Human polyomavirus 6 (HPyV6) and HPyV7 are commonly found on human skin. We have determined the X-ray structures of their major capsid protein, VP1, at resolutions of 1.8 and 1.7 Å, respectively. In polyomaviruses, VP1 commonly determines antigenicity as well as cell-surface receptor specificity, and the protein is therefore linked to attachment, tropism, and ultimately, viral pathogenicity. The structures of HPyV6 and HPyV7 VP1 reveal uniquely elongated loops that cover the bulk of the outer virion surfaces, obstructing a groove that binds sialylated glycan receptors in many other polyomaviruses. In support of this structural observation, interactions of VP1 with α2,3- and α2,6-linked sialic acids could not be detected in solution by nuclear magnetic resonance spectroscopy. Single-cell binding studies indicate that sialylated glycans are likely not required for initial attachment to cultured human cells. Our findings establish distinct antigenic properties of HPyV6 and HPyV7 capsids and indicate that these two viruses engage nonsialylated receptors.
Importance: Eleven new human polyomaviruses, including the skin viruses HPyV6 and HPyV7, have been identified during the last decade. In contrast to better-studied polyomaviruses, the routes of infection, cell tropism, and entry pathways of many of these new viruses remain largely mysterious. Our high-resolution X-ray structures of major capsid proteins VP1 from HPyV6 and from HPyV7 reveal critical differences in surface morphology from those of all other known polyomavirus structures. A groove that engages specific sialic acid-containing glycan receptors in related polyomaviruses is obstructed, and VP1 of HPyV6 and HPyV7 does not interact with sialylated compounds in solution or on cultured human cells. A comprehensive comparison with other structurally characterized polyomavirus VP1 proteins enhances our understanding of molecular determinants that underlie receptor specificity, antigenicity, and, ultimately, pathogenicity within the polyomavirus family and highlight the need for structure-based analysis to better define phylogenetic relationships within the growing polyomavirus family and perhaps also for other viruses.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
Trichodysplasia spinulosa-Associated Polyomavirus Uses a Displaced Binding Site on VP1 to Engage Sialylated Glycolipids.PLoS Pathog. 2015 Aug 24;11(8):e1005112. doi: 10.1371/journal.ppat.1005112. eCollection 2015 Aug. PLoS Pathog. 2015. PMID: 26302170 Free PMC article.
-
Structural Basis and Evolution of Glycan Receptor Specificities within the Polyomavirus Family.mBio. 2020 Jul 28;11(4):e00745-20. doi: 10.1128/mBio.00745-20. mBio. 2020. PMID: 32723915 Free PMC article.
-
Seroprevalence rates of HPyV6, HPyV7, TSPyV, HPyV9, MWPyV and KIPyV polyomaviruses among the healthy blood donors.J Med Virol. 2016 Jul;88(7):1254-61. doi: 10.1002/jmv.24440. Epub 2015 Dec 15. J Med Virol. 2016. PMID: 26630080
-
Genome analysis of the new human polyomaviruses.Rev Med Virol. 2012 Nov;22(6):354-77. doi: 10.1002/rmv.1711. Epub 2012 Mar 28. Rev Med Virol. 2012. PMID: 22461085 Review.
-
The Polyomaviridae: Contributions of virus structure to our understanding of virus receptors and infectious entry.Virology. 2009 Feb 20;384(2):389-99. doi: 10.1016/j.virol.2008.12.021. Epub 2009 Jan 21. Virology. 2009. PMID: 19157478 Free PMC article. Review.
Cited by
-
EXTL3-interacting endometriosis-specific serum factors induce colony formation of endometrial stromal cells.Sci Rep. 2019 Aug 29;9(1):12562. doi: 10.1038/s41598-019-48840-8. Sci Rep. 2019. PMID: 31467315 Free PMC article.
-
Taking the Scenic Route: Polyomaviruses Utilize Multiple Pathways to Reach the Same Destination.Viruses. 2020 Oct 15;12(10):1168. doi: 10.3390/v12101168. Viruses. 2020. PMID: 33076363 Free PMC article. Review.
-
In Vitro and In Vivo Models for the Study of Human Polyomavirus Infection.Viruses. 2016 Oct 22;8(10):292. doi: 10.3390/v8100292. Viruses. 2016. PMID: 27782080 Free PMC article. Review.
-
The Biology and Clinical Features of Cutaneous Polyomaviruses.J Invest Dermatol. 2019 Feb;139(2):285-292. doi: 10.1016/j.jid.2018.09.013. Epub 2018 Nov 22. J Invest Dermatol. 2019. PMID: 30470393 Free PMC article. Review.
-
Trichodysplasia spinulosa-Associated Polyomavirus Uses a Displaced Binding Site on VP1 to Engage Sialylated Glycolipids.PLoS Pathog. 2015 Aug 24;11(8):e1005112. doi: 10.1371/journal.ppat.1005112. eCollection 2015 Aug. PLoS Pathog. 2015. PMID: 26302170 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
