

Quantitative determinants of aerobic glycolysis identify flux through the enzyme GAPDH as a limiting step
- PMID: 25009227
- PMCID: PMC4118620
- DOI: 10.7554/eLife.03342
Quantitative determinants of aerobic glycolysis identify flux through the enzyme GAPDH as a limiting step
Abstract
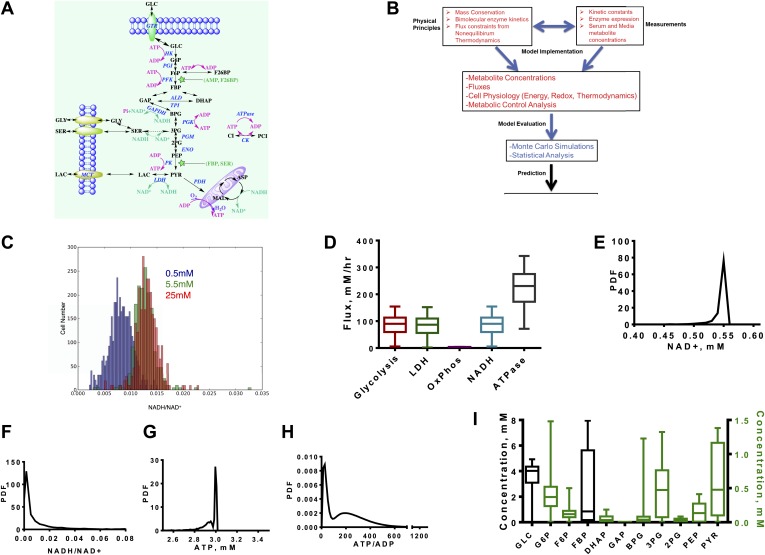
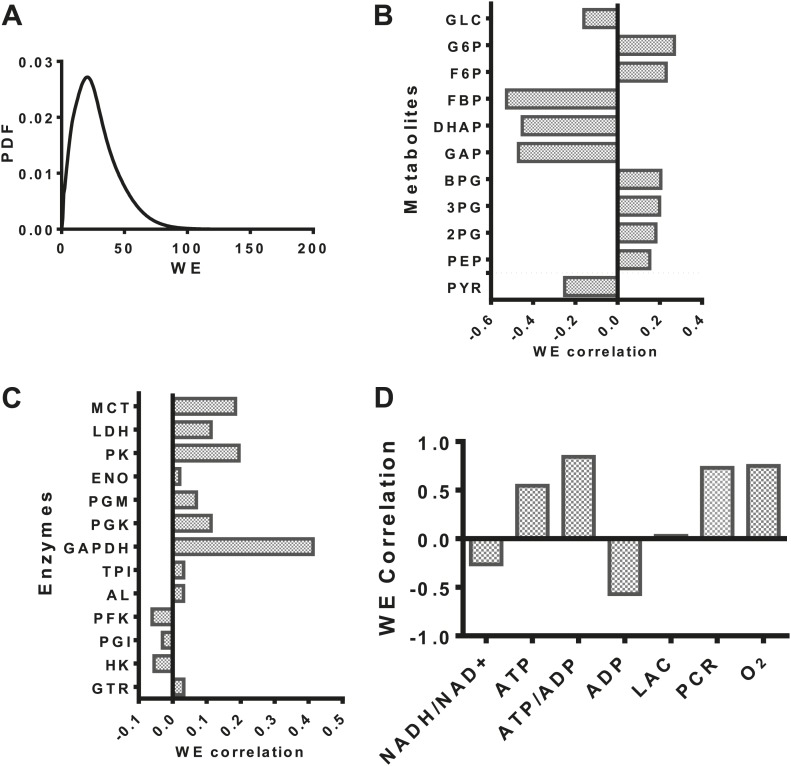
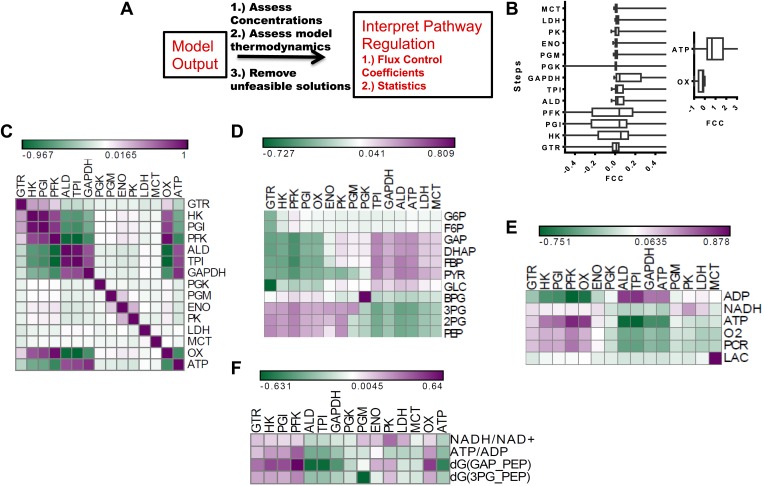
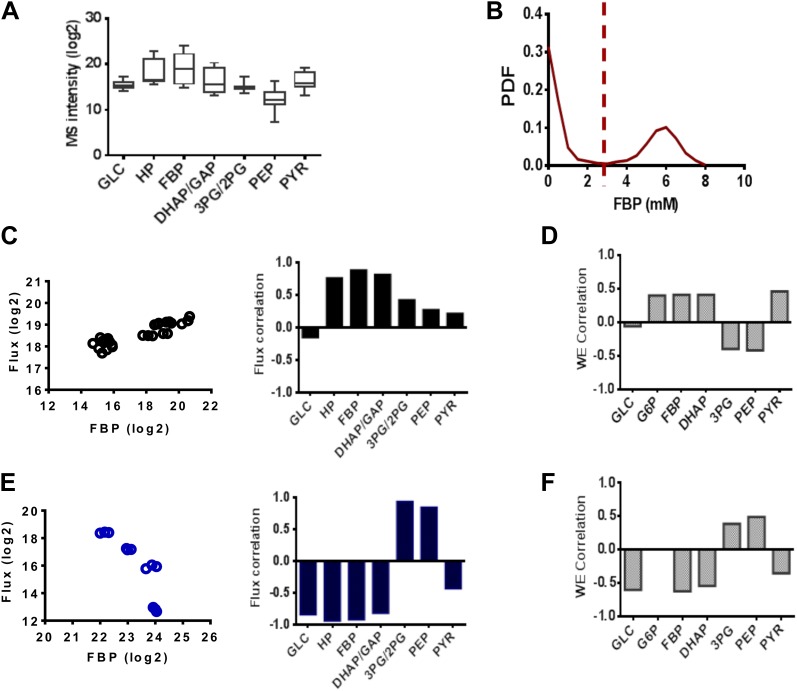
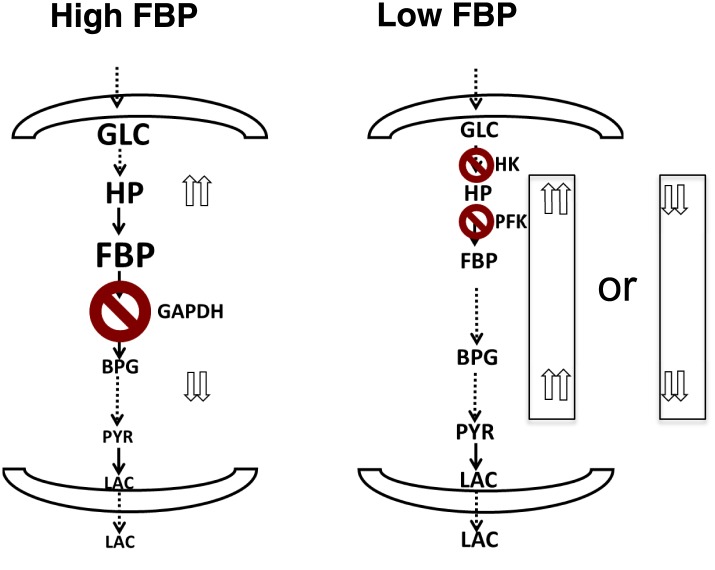
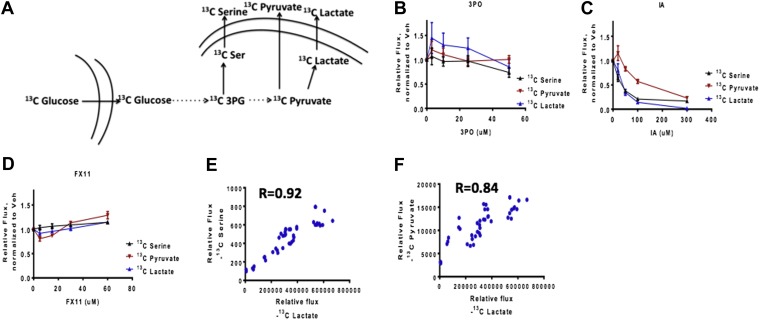
Aerobic glycolysis or the Warburg Effect (WE) is characterized by the increased metabolism of glucose to lactate. It remains unknown what quantitative changes to the activity of metabolism are necessary and sufficient for this phenotype. We developed a computational model of glycolysis and an integrated analysis using metabolic control analysis (MCA), metabolomics data, and statistical simulations. We identified and confirmed a novel mode of regulation specific to aerobic glycolysis where flux through GAPDH, the enzyme separating lower and upper glycolysis, is the rate-limiting step in the pathway and the levels of fructose (1,6) bisphosphate (FBP), are predictive of the rate and control points in glycolysis. Strikingly, negative flux control was found and confirmed for several steps thought to be rate-limiting in glycolysis. Together, these findings enumerate the biochemical determinants of the WE and suggest strategies for identifying the contexts in which agents that target glycolysis might be most effective.
Keywords: biochemistry; glucose; glycolysis; human; human biology; mass spectrometry; mathematical modeling; medicine; metabolism; metabolomics.
Conflict of interest statement
JWL: A patent related to this work has been filed. US Provisional Patent Appln. No. 61/908,953.
The other authors declare that no competing interests exist.
Figures

Similar articles
-
Evolved resistance to partial GAPDH inhibition results in loss of the Warburg effect and in a different state of glycolysis.J Biol Chem. 2020 Jan 3;295(1):111-124. doi: 10.1074/jbc.RA119.010903. Epub 2019 Nov 20. J Biol Chem. 2020. PMID: 31748414 Free PMC article.
-
Determining the quantitative relationship between glycolysis and GAPDH in cancer cells exhibiting the Warburg effect.J Biol Chem. 2021 Jan-Jun;296:100369. doi: 10.1016/j.jbc.2021.100369. Epub 2021 Feb 3. J Biol Chem. 2021. PMID: 33545174 Free PMC article.
-
Natural product 1,2,3,4,6-penta-O-galloyl-β-D-glucopyranose is a reversible inhibitor of glyceraldehyde 3-phosphate dehydrogenase.Acta Pharmacol Sin. 2022 Feb;43(2):470-482. doi: 10.1038/s41401-021-00653-0. Epub 2021 Apr 13. Acta Pharmacol Sin. 2022. PMID: 33850276 Free PMC article.
-
Age-dependent metabolic dysregulation in cancer and Alzheimer's disease.Biogerontology. 2014 Dec;15(6):559-77. doi: 10.1007/s10522-014-9534-z. Epub 2014 Oct 11. Biogerontology. 2014. PMID: 25305052 Review.
-
Cancer metabolism: what we can learn from proteomic analysis by mass spectrometry.Cancer Genomics Proteomics. 2012 Nov;9(6):373-81. Cancer Genomics Proteomics. 2012. PMID: 23162076 Free PMC article. Review.
Cited by
-
Organization of enzyme concentration across the metabolic network in cancer cells.PLoS One. 2015 Jan 26;10(1):e0117131. doi: 10.1371/journal.pone.0117131. eCollection 2015. PLoS One. 2015. PMID: 25621879 Free PMC article.
-
Long noncoding RNA CASC7 is a novel regulator of glycolysis in oesophageal cancer via a miR-143-3p-mediated HK2 signalling pathway.Cell Death Discov. 2022 Apr 26;8(1):231. doi: 10.1038/s41420-022-01028-y. Cell Death Discov. 2022. PMID: 35474307 Free PMC article.
-
Glucose Metabolites Exert Opposing Roles in Tumor Chemoresistance.Front Oncol. 2019 Nov 21;9:1282. doi: 10.3389/fonc.2019.01282. eCollection 2019. Front Oncol. 2019. PMID: 31824857 Free PMC article.
-
Multiple Myeloma and Fatty Acid Metabolism.JBMR Plus. 2019 Feb 20;3(3):e10173. doi: 10.1002/jbm4.10173. eCollection 2019 Mar. JBMR Plus. 2019. PMID: 30918920 Free PMC article. Review.
-
Uncoupled glycerol-3-phosphate shuttle in kidney cancer reveals that cytosolic GPD is essential to support lipid synthesis.Mol Cell. 2023 Apr 20;83(8):1340-1349.e7. doi: 10.1016/j.molcel.2023.03.023. Mol Cell. 2023. PMID: 37084714 Free PMC article.
References
-
- Anastasiou D, Poulogiannis G, Asara JM, Boxer MB, Jiang JK, Shen M, Bellinger G, Sasaki AT, Locasale JW, Auld DS, Thomas CJ, Vander Heiden MG, Cantley LC. 2011. Inhibition of pyruvate kinase M2 by reactive oxygen species contributes to cellular antioxidant responses. Science 334:1278–1283. doi: 10.1126/science.1211485 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
