

Bacterial cell wall. MurJ is the flippase of lipid-linked precursors for peptidoglycan biogenesis
- PMID: 25013077
- PMCID: PMC4163187
- DOI: 10.1126/science.1254522
Bacterial cell wall. MurJ is the flippase of lipid-linked precursors for peptidoglycan biogenesis
Abstract
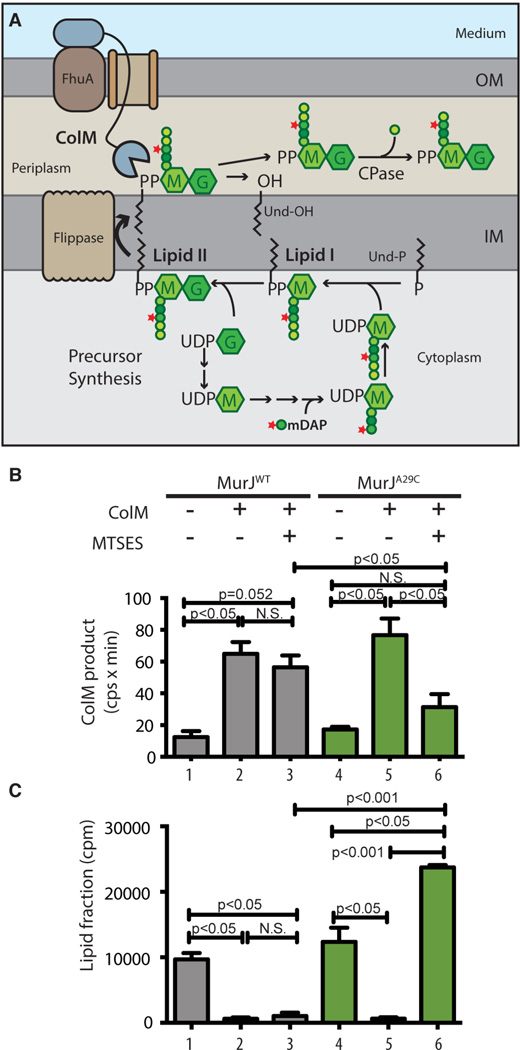
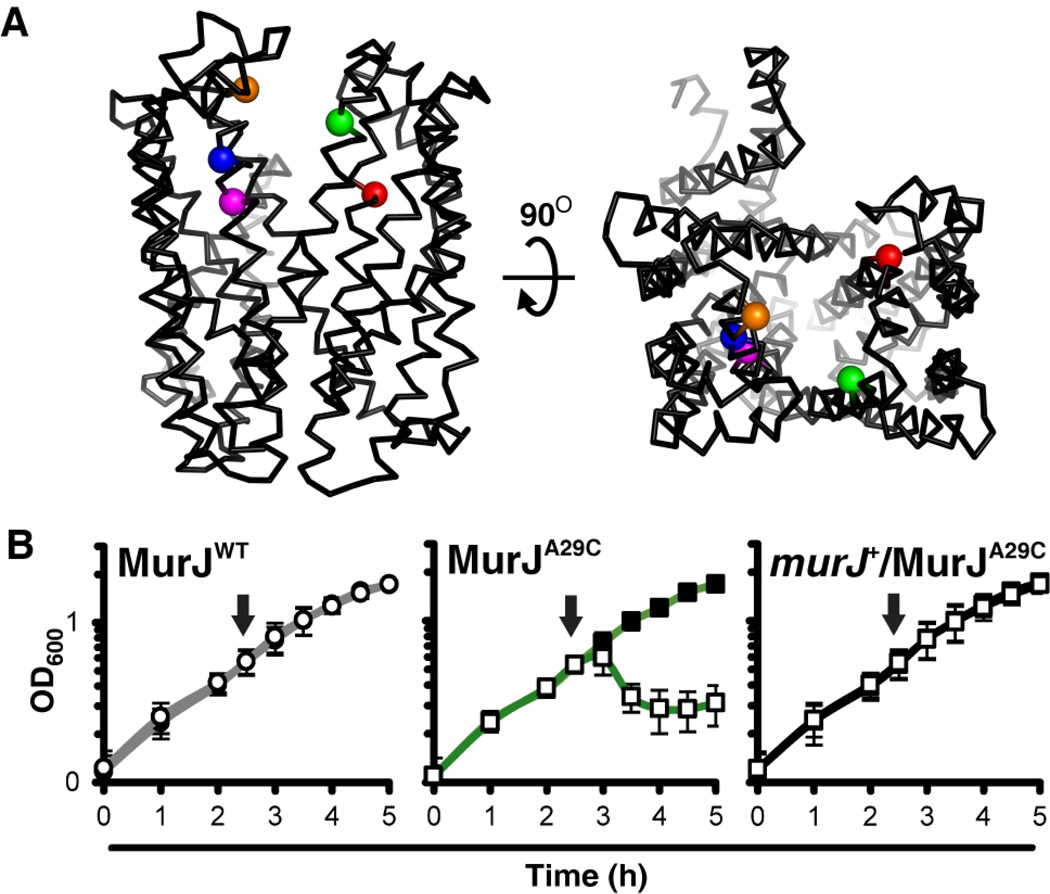
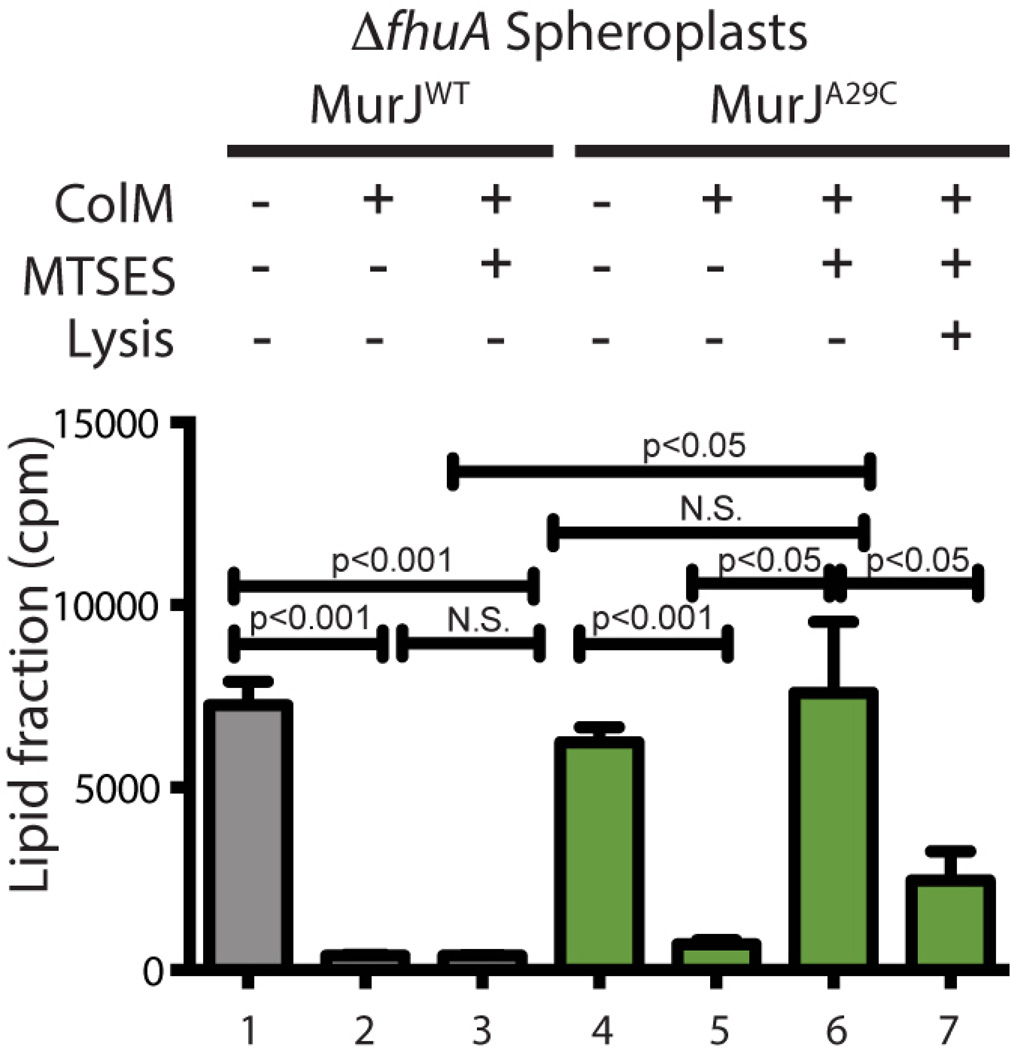
Peptidoglycan (PG) is a polysaccharide matrix that protects bacteria from osmotic lysis. Inhibition of its biogenesis is a proven strategy for killing bacteria with antibiotics. The assembly of PG requires disaccharide-pentapeptide building blocks attached to a polyisoprene lipid carrier called lipid II. Although the stages of lipid II synthesis are known, the identity of the essential flippase that translocates it across the cytoplasmic membrane for PG polymerization is unclear. We developed an assay for lipid II flippase activity and used a chemical genetic strategy to rapidly and specifically block flippase function. We combined these approaches to demonstrate that MurJ is the lipid II flippase in Escherichia coli.
Copyright © 2014, American Association for the Advancement of Science.
Figures
Comment in
-
Microbiology. A flipping cell wall ferry.Science. 2014 Jul 11;345(6193):139-40. doi: 10.1126/science.1256585. Science. 2014. PMID: 25013047 No abstract available.
-
Bacterial physiology: Flipping out over MurJ.Nat Rev Microbiol. 2014 Sep;12(9):595. doi: 10.1038/nrmicro3328. Epub 2014 Jul 21. Nat Rev Microbiol. 2014. PMID: 25043163 No abstract available.
References
-
- Bouhss A, Trunkfield AE, Bugg TD, Mengin-Lecreulx D. The biosynthesis of peptidoglycan lipid-linked intermediates. FEMS Microbiol Rev. 2008 Mar;32:208. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
