

Phospho-dependent and phospho-independent interactions of the helicase UPF1 with the NMD factors SMG5-SMG7 and SMG6
- PMID: 25013172
- PMCID: PMC4132714
- DOI: 10.1093/nar/gku578
Phospho-dependent and phospho-independent interactions of the helicase UPF1 with the NMD factors SMG5-SMG7 and SMG6
Abstract
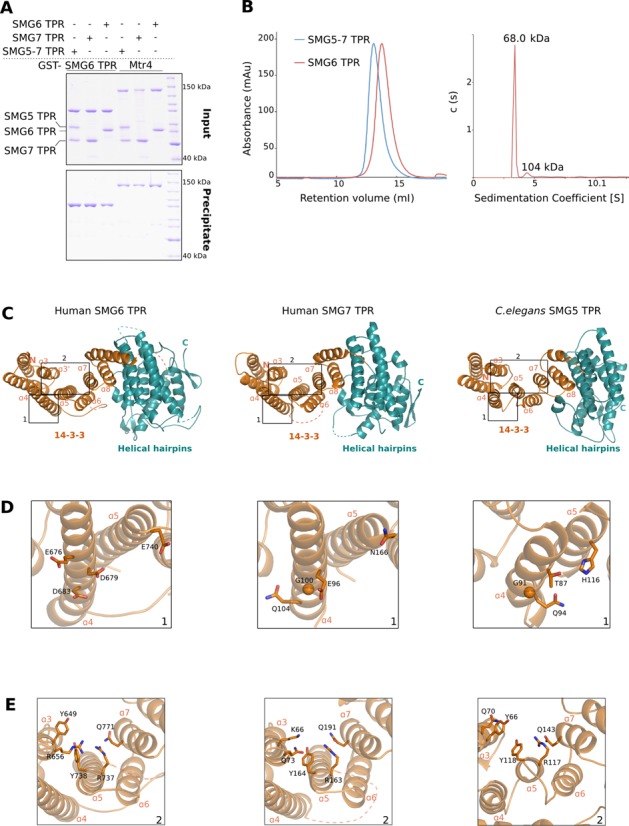
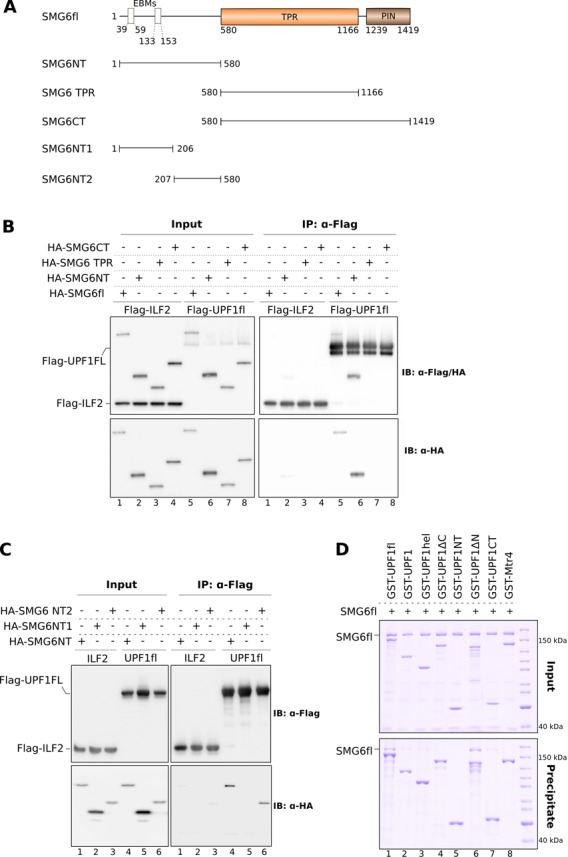
Nonsense-mediated mRNA decay (NMD) is a eukaryotic surveillance pathway that recognizes mRNAs with premature stop codons and targets them for rapid degradation. Evidence from previous studies has converged on UPF1 as the central NMD factor. In human cells, the SMG1 kinase phosphorylates UPF1 at the N-terminal and C-terminal tails, in turn allowing the recruitment of the NMD factors SMG5, SMG6 and SMG7. To understand the molecular mechanisms, we recapitulated these steps of NMD in vitro using purified components. We find that a short C-terminal segment of phosphorylated UPF1 containing the last two Ser-Gln motifs is recognized by the heterodimer of SMG5 and SMG7 14-3-3-like proteins. In contrast, the SMG6 14-3-3-like domain is a monomer. The crystal structure indicates that the phosphoserine binding site of the SMG6 14-3-3-like domain is similar to that of SMG5 and can mediate a weak phospho-dependent interaction with UPF1. The dominant SMG6-UPF1 interaction is mediated by a low-complexity region bordering the 14-3-3-like domain of SMG6 and by the helicase domain and C-terminal tail of UPF1. This interaction is phosphorylation independent. Our study demonstrates that SMG5-SMG7 and SMG6 exhibit different and non-overlapping modes of UPF1 recognition, thus pointing at distinguished roles in integrating the complex NMD interaction network.
© The Author(s) 2014. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures



References
-
- Bhuvanagiri M., Schlitter A.M., Hentze M.W., Kulozik A.E. NMD: RNA biology meets human genetic medicine. Biochem. J. 2010;430:365–377. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
