

Identification of a functional interaction of HMGB1 with Receptor for Advanced Glycation End-products in a model of neuropathic pain
- PMID: 25014009
- PMCID: PMC4560334
- DOI: 10.1016/j.bbi.2014.06.199
Identification of a functional interaction of HMGB1 with Receptor for Advanced Glycation End-products in a model of neuropathic pain
Abstract
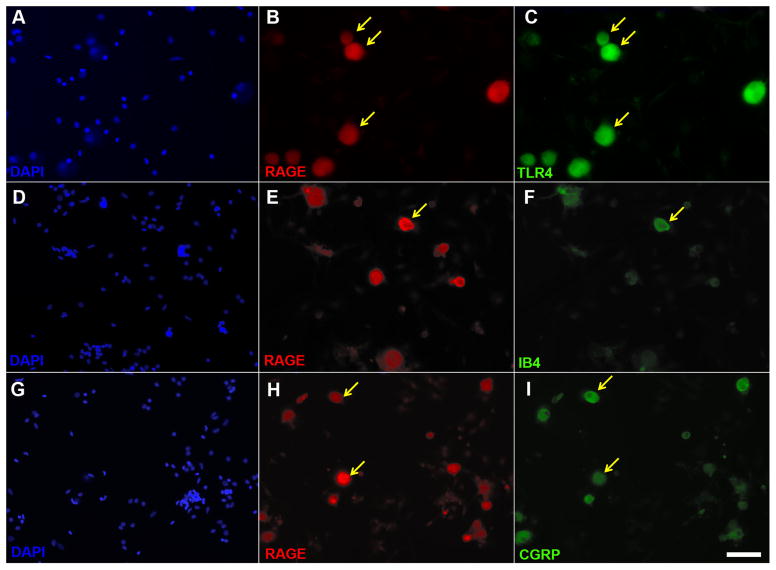
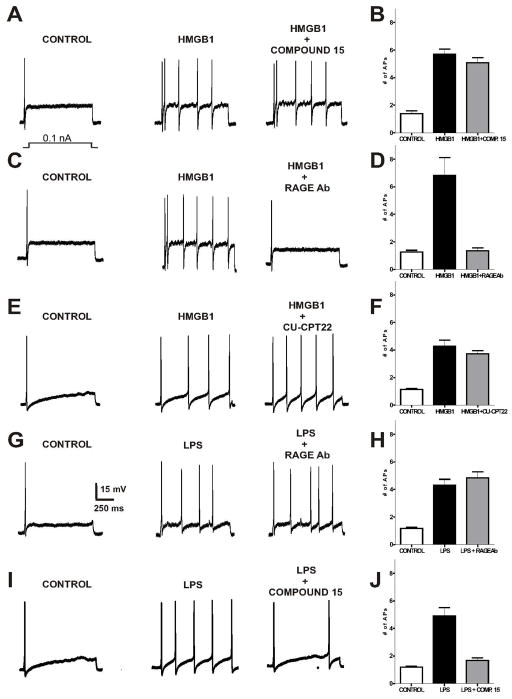
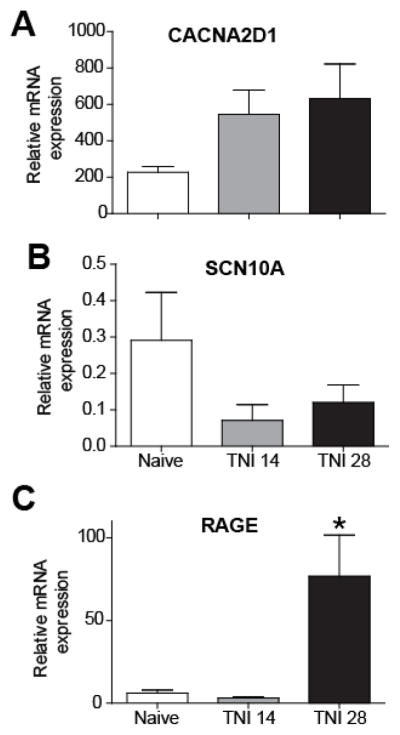
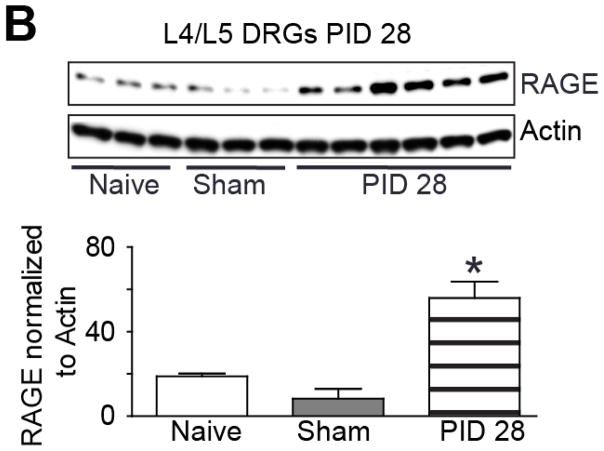
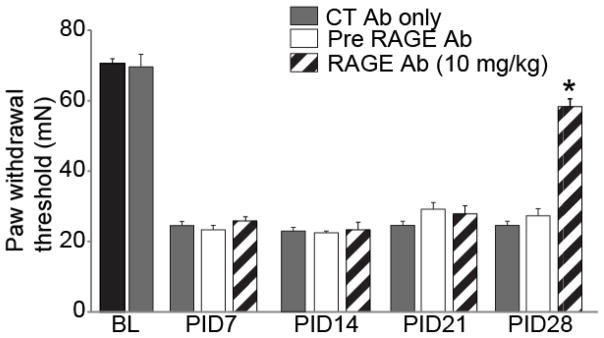
Recent studies indicate that the release of high mobility group box 1 (HMGB1) following nerve injury may play a central role in the pathogenesis of neuropathic pain. HMGB1 is known to influence cellular responses within the nervous system via two distinct receptor families; the Receptor for Advanced Glycation End-products (RAGE) and Toll-like receptors (TLRs). The degree to which HMGB1 activates a receptor is thought to be dependent upon the oxidative state of the ligand, resulting in the functional isoforms of all-thiol HMGB1 (at-HMGB1) acting through RAGE, and disufide HMGB1 (ds-HMGB1) interacting with TLR4. Though it is known that dorsal root ganglia (DRG) sensory neurons exposed to HMGB1 and TLR4 agonists can influence excitation, the degree to which at-HMGB1 signaling through neuronal RAGE contributes to neuropathic pain is unknown. Here we demonstrate that at-HMGB1 activation of nociceptive neurons is dependent on RAGE and not TLR4. To distinguish the possible role of RAGE on neuropathic pain, we characterized the changes in RAGE mRNA expression up to one month after tibial nerve injury (TNI). RAGE mRNA expression in lumbar dorsal root ganglion (DRG) is substantially increased by post-injury day (PID) 28 when compared with sham injured rodents. Protein expression at PID28 confirms this injury-induced event in the DRG. Moreover, a single exposure to monoclonal antibody to RAGE (RAGE Ab) failed to abrogate pain behavior at PID 7, 14 and 21. However, RAGE Ab administration produced reversal of mechanical hyperalgesia on PID28. Thus, at-HMGB1 activation through RAGE may be responsible for sensory neuron sensitization and mechanical hyperalgesia associated with chronic neuropathic pain states.
Keywords: Alarmin; DAMP; DRG; Dorsal root ganglia; HMGB1; Pain; RAGE; TLR4.
Copyright © 2014 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
-
- Calvo M, Dawes JM, Bennett DL. The role of the immune system in the generation of neuropathic pain. Lancet Neurol. 2012;11:629–42. - PubMed
-
- Bianchi ME. DAMPs, PAMPs and alarmins: all we need to know about danger. J Leukoc Biol. 2007;81:1–5. - PubMed
-
- Czura CJ, Wang H, Tracey KJ. Dual roles for HMGB1: DNA binding and cytokine. J Endotoxin Res. 2001;7:315–21. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
