

Gβ1γ2 activates phospholipase A2-dependent Golgi membrane tubule formation
- PMID: 25019068
- PMCID: PMC4091998
- DOI: 10.3389/fcell.2014.00004
Gβ1γ2 activates phospholipase A2-dependent Golgi membrane tubule formation
Abstract
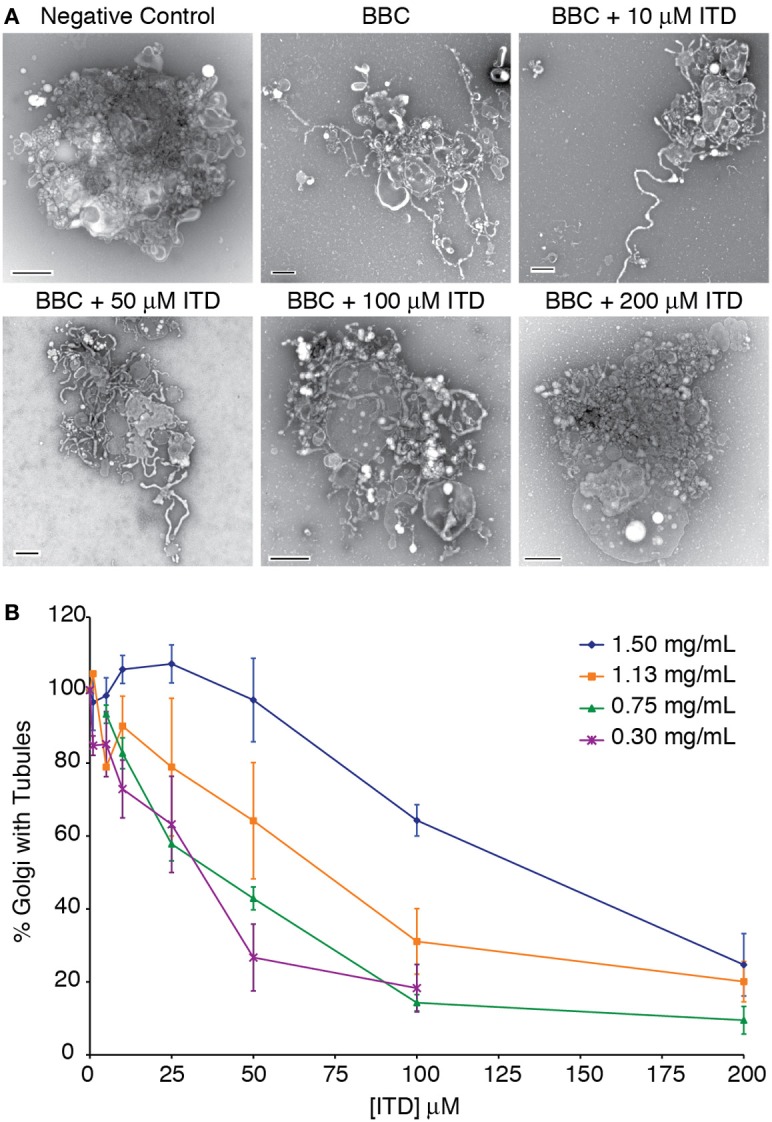
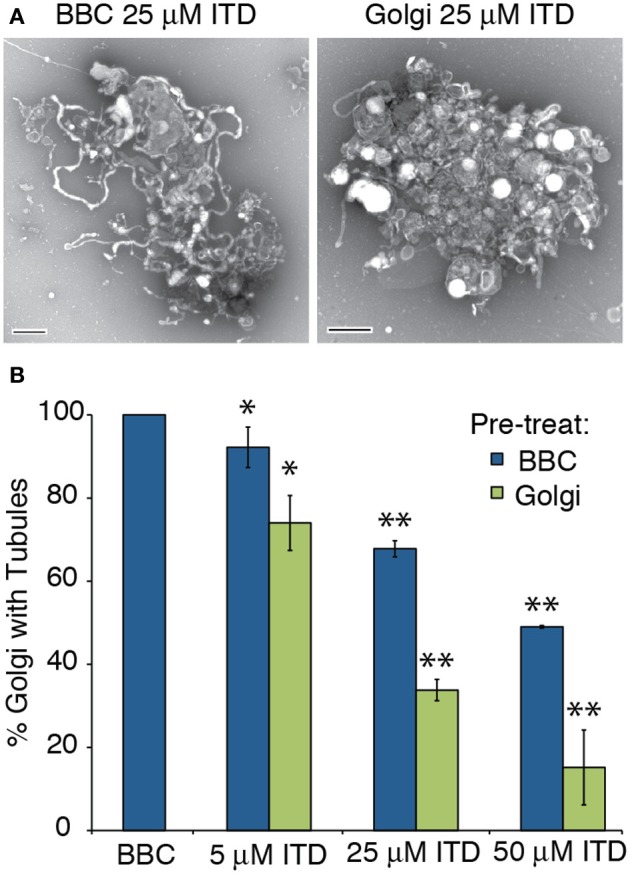
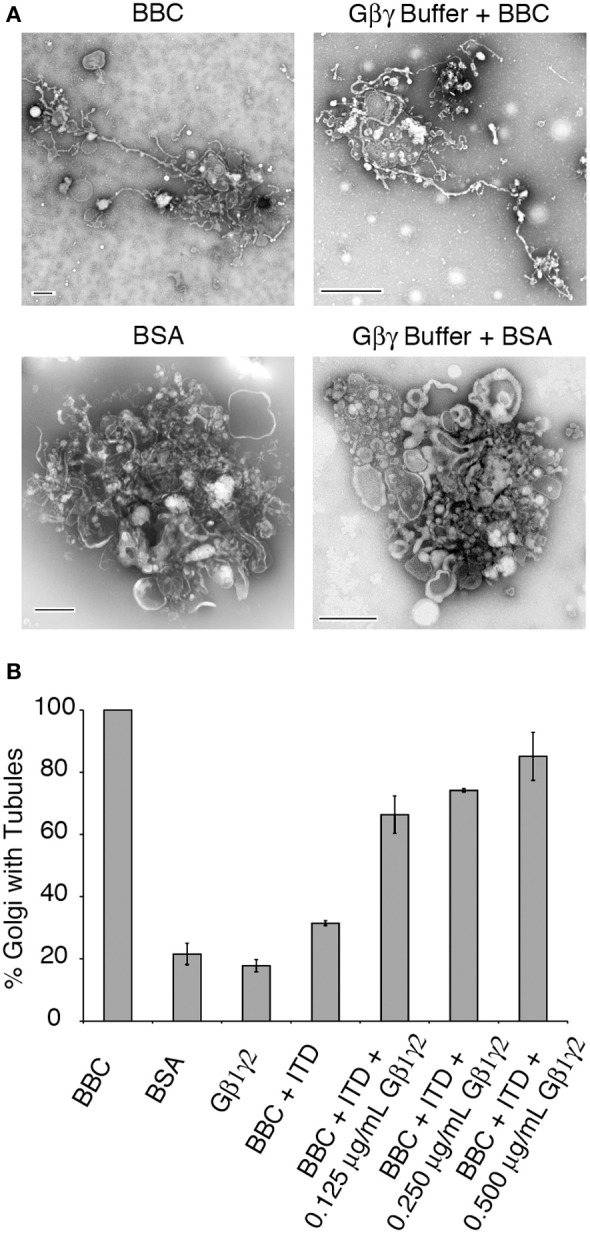
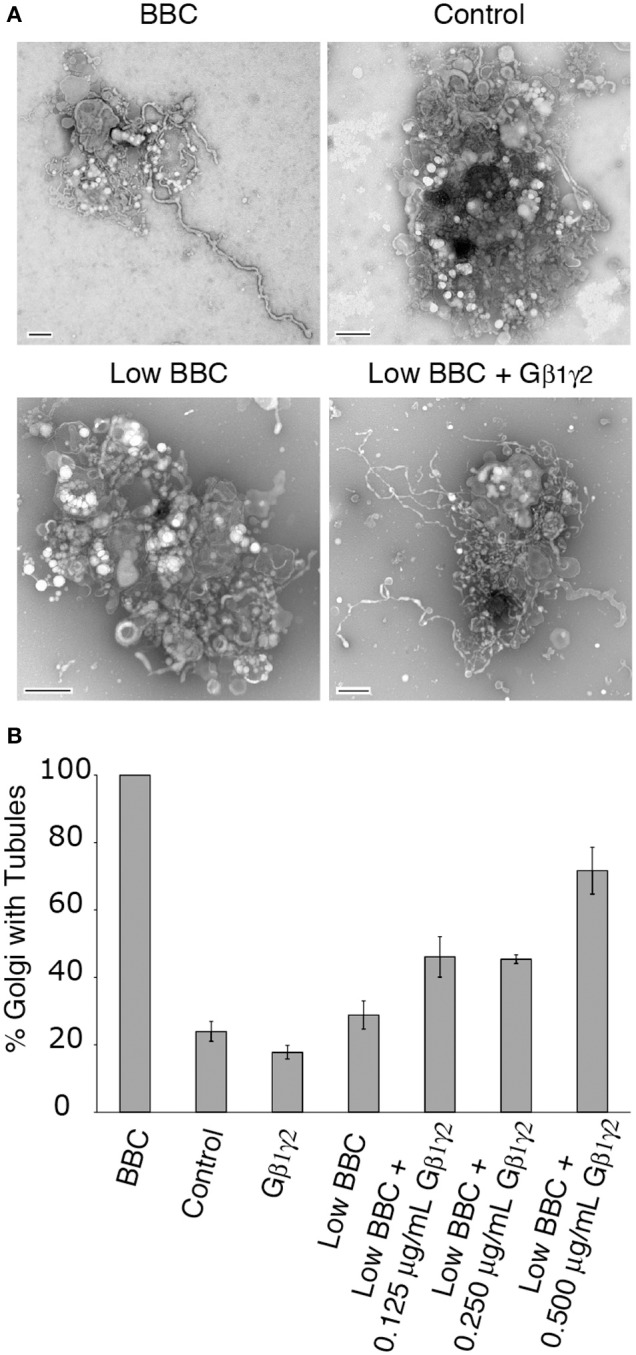
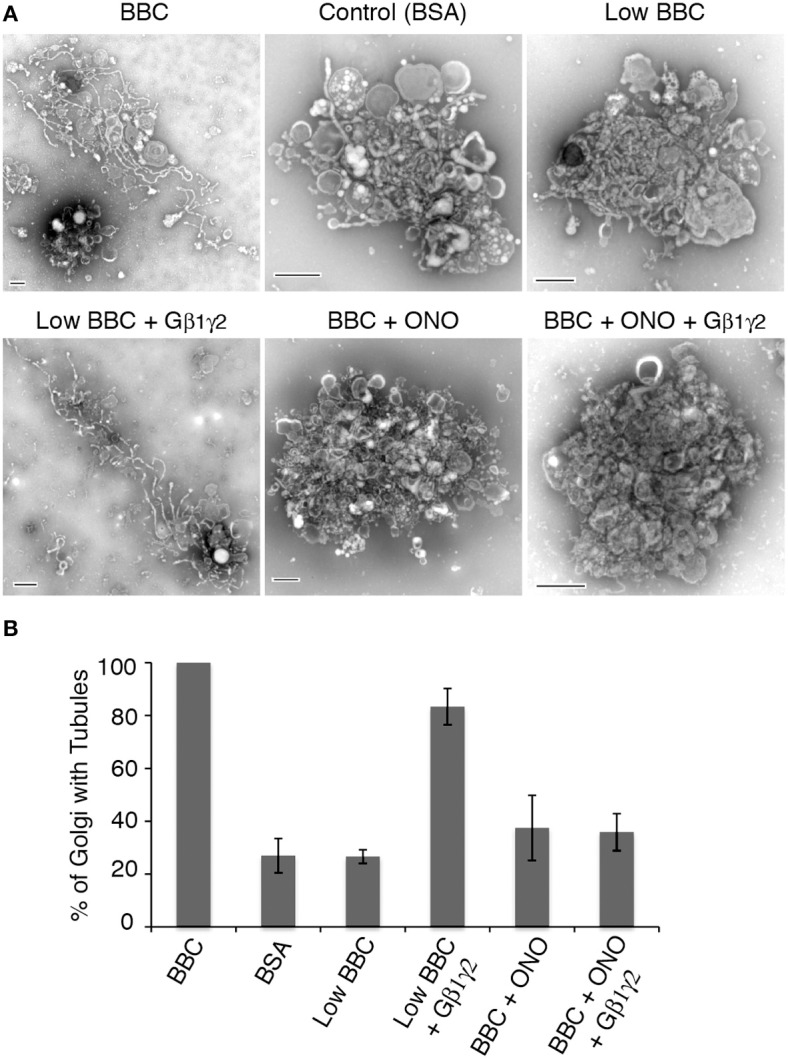
Heterotrimeric G proteins transduce the ligand binding of transmembrane G protein coupled receptors into a variety of intracellular signaling pathways. Recently, heterotrimeric Gβγ subunit signaling at the Golgi complex has been shown to regulate the formation of vesicular transport carriers that deliver cargo from the Golgi to the plasma membrane. In addition to vesicles, membrane tubules have also been shown to mediate export from the Golgi complex, which requires the activity of cytoplasmic phospholipase A2 (PLA2) enzyme activity. Through the use of an in vitro reconstitution assay with isolated Golgi complexes, we provide evidence that Gβ1γ2 signaling also stimulates Golgi membrane tubule formation. In addition, we show that an inhibitor of Gβγ activation of PLA2 enzymes inhibits in vitro Golgi membrane tubule formation. Additionally, purified Gβγ protein stimulates membrane tubules in the presence of low (sub-threshold) cytosol concentrations. Importantly, this Gβγ stimulation of Golgi membrane tubule formation was inhibited by treatment with the PLA2 antagonist ONO-RS-082. These studies indicate that Gβ1γ2 signaling activates PLA2 enzymes required for Golgi membrane tubule formation, thus establishing a new layer of regulation for this process.
Keywords: Golgi complex; Gβγ; PLA2; heterotrimeric G proteins; membrane tubules; phospholipids.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources