

Acute brain slice methods for adult and aging animals: application of targeted patch clamp analysis and optogenetics
- PMID: 25023312
- PMCID: PMC4219416
- DOI: 10.1007/978-1-4939-1096-0_14
Acute brain slice methods for adult and aging animals: application of targeted patch clamp analysis and optogenetics
Abstract
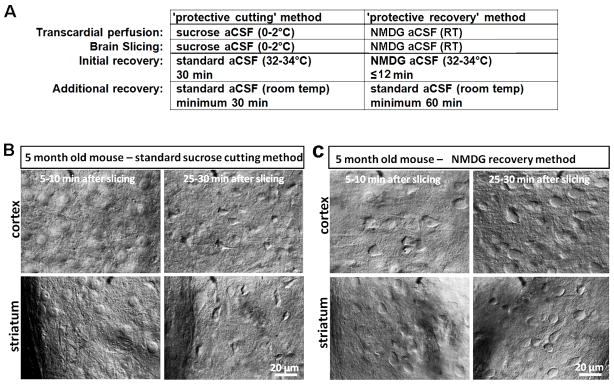
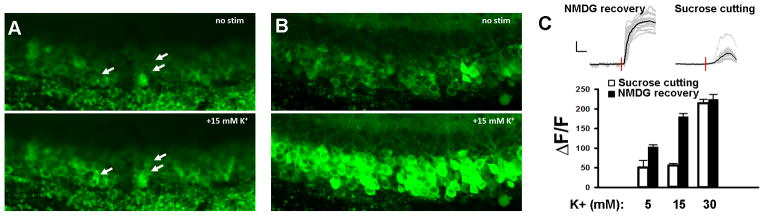
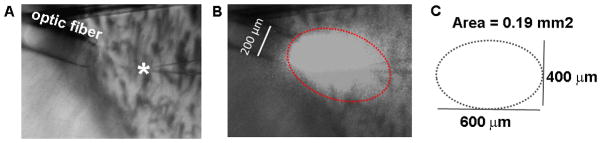
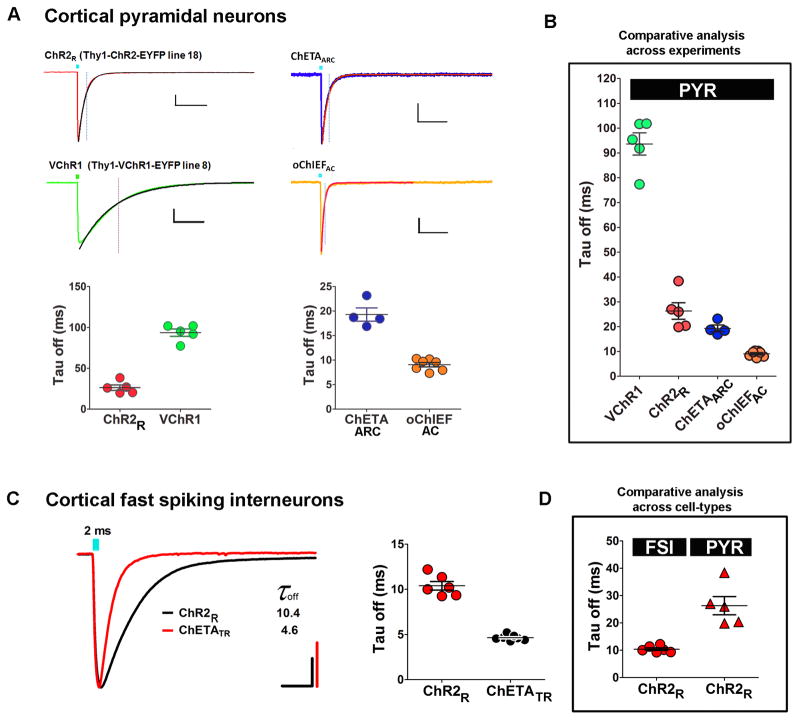
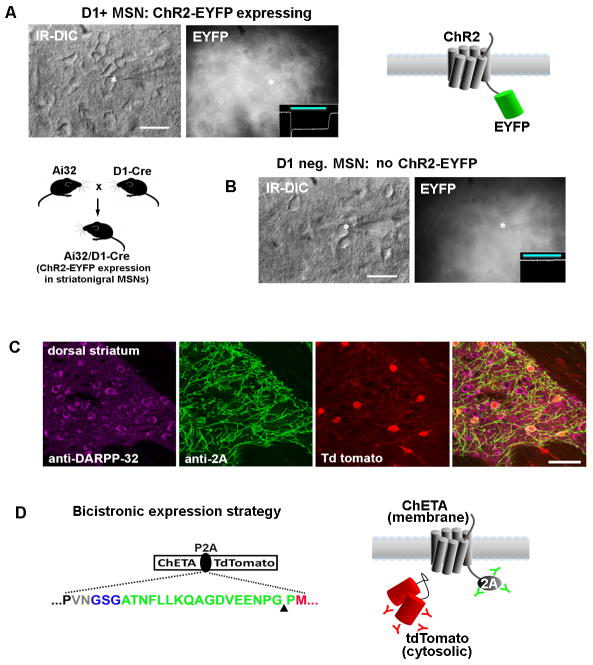
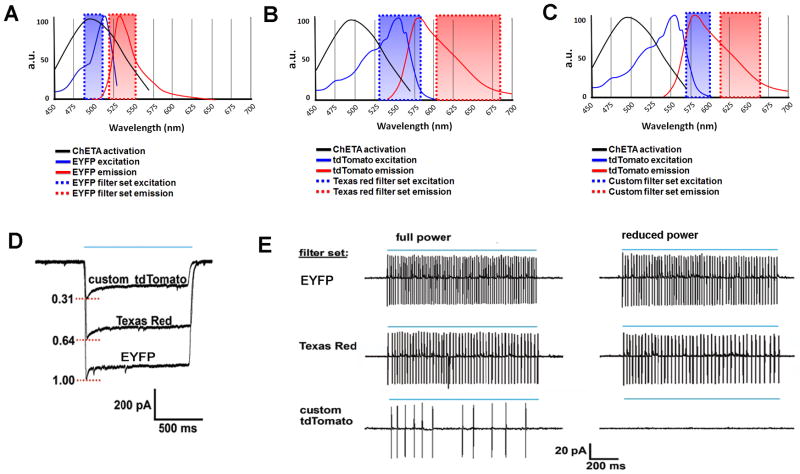
The development of the living acute brain slice preparation for analyzing synaptic function roughly a half century ago was a pivotal achievement that greatly influenced the landscape of modern neuroscience. Indeed, many neuroscientists regard brain slices as the gold-standard model system for detailed cellular, molecular, and circuitry level analysis and perturbation of neuronal function. A critical limitation of this model system is the difficulty in preparing slices from adult and aging animals, and over the past several decades few substantial methodological improvements have emerged to facilitate patch clamp analysis in the mature adult stage. In this chapter we describe a robust and practical protocol for preparing brain slices from mature adult mice that are suitable for patch clamp analysis. This method reduces swelling and damage in superficial layers of the slices and improves the success rate for targeted patch clamp recordings, including recordings from fluorescently labeled populations in slices derived from transgenic mice. This adult brain slice method is suitable for diverse experimental applications, including both monitoring and manipulating neuronal activity with genetically encoded calcium indicators and optogenetic actuators, respectively. We describe the application of this adult brain slice platform and associated methods for screening kinetic properties of Channelrhodopsin (ChR) variants expressed in genetically defined neuronal subtypes.
Figures

Similar articles
-
Single or Double Patch-Clamp Recordings In Ex Vivo Slice Preparation: Functional Connectivity, Synapse Dynamics, and Optogenetics.Methods Mol Biol. 2021;2188:285-309. doi: 10.1007/978-1-0716-0818-0_15. Methods Mol Biol. 2021. PMID: 33119858
-
Studying Neuronal Function Ex Vivo Using Optogenetic Stimulation and Patch Clamp.Methods Mol Biol. 2020;2173:1-20. doi: 10.1007/978-1-0716-0755-8_1. Methods Mol Biol. 2020. PMID: 32651907
-
Optogenetics to Interrogate Neuron-Glia Interactions in Pups and Adults.Methods Mol Biol. 2021;2191:135-149. doi: 10.1007/978-1-0716-0830-2_9. Methods Mol Biol. 2021. PMID: 32865743
-
Studying Synaptic Connectivity and Strength with Optogenetics and Patch-Clamp Electrophysiology.Int J Mol Sci. 2022 Oct 1;23(19):11612. doi: 10.3390/ijms231911612. Int J Mol Sci. 2022. PMID: 36232917 Free PMC article. Review.
-
Recent advances in cellular optogenetics for photomedicine.Adv Drug Deliv Rev. 2022 Sep;188:114457. doi: 10.1016/j.addr.2022.114457. Epub 2022 Jul 16. Adv Drug Deliv Rev. 2022. PMID: 35843507 Review.
Cited by
-
TRPV1-mediated presynaptic transmission in basolateral amygdala contributes to visceral hypersensitivity in adult rats with neonatal maternal deprivation.Sci Rep. 2016 Jul 1;6:29026. doi: 10.1038/srep29026. Sci Rep. 2016. PMID: 27364923 Free PMC article.
-
Heterozygous expression of a Kcnt1 gain-of-function variant has differential effects on somatostatin- and parvalbumin-expressing cortical GABAergic neurons.Elife. 2024 Oct 11;13:RP92915. doi: 10.7554/eLife.92915. Elife. 2024. PMID: 39392867 Free PMC article.
-
Effect of brain acidification on depression-related behaviors in diabetes mellitus.Front Psychiatry. 2023 Nov 29;14:1277097. doi: 10.3389/fpsyt.2023.1277097. eCollection 2023. Front Psychiatry. 2023. PMID: 38094594 Free PMC article.
-
Transcranial direct current stimulation relieves visceral hypersensitivity via normalizing GluN2B expression and neural activity in anterior cingulate cortex.J Neurophysiol. 2021 May 1;125(5):1787-1797. doi: 10.1152/jn.00025.2021. Epub 2021 Mar 24. J Neurophysiol. 2021. PMID: 33760644 Free PMC article.
-
Chronically dysregulated corticosterone impairs dopaminergic transmission in the dorsomedial striatum by sex-divergent mechanisms.Neuropsychopharmacology. 2023 Aug;48(9):1328-1337. doi: 10.1038/s41386-023-01551-1. Epub 2023 Feb 21. Neuropsychopharmacology. 2023. PMID: 36810463 Free PMC article.
References
-
- Moyer JR, Jr, Brown TH. Methods for whole-cell recording from visually preselected neurons of perirhinal cortex in brain slices from young and aging rats. J Neurosci Meth. 1998;86:35–54. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
