

Bridging integrator 1 (BIN1) protein expression increases in the Alzheimer's disease brain and correlates with neurofibrillary tangle pathology
- PMID: 25024306
- PMCID: PMC4198456
- DOI: 10.3233/JAD-132450
Bridging integrator 1 (BIN1) protein expression increases in the Alzheimer's disease brain and correlates with neurofibrillary tangle pathology
Abstract
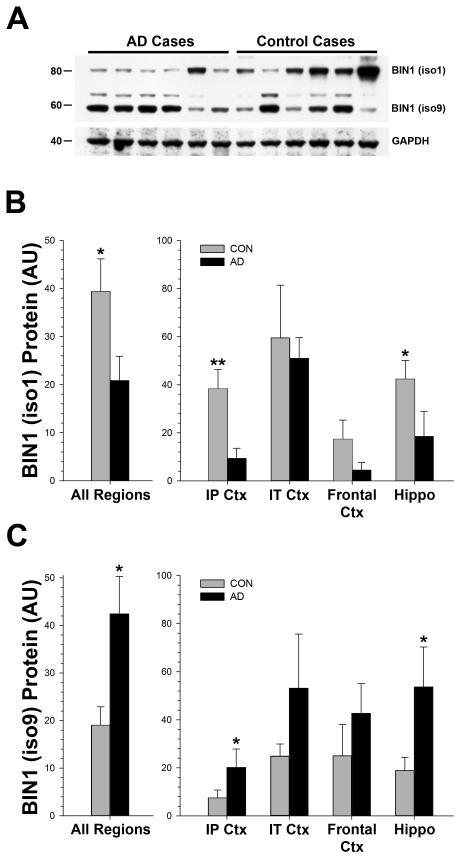
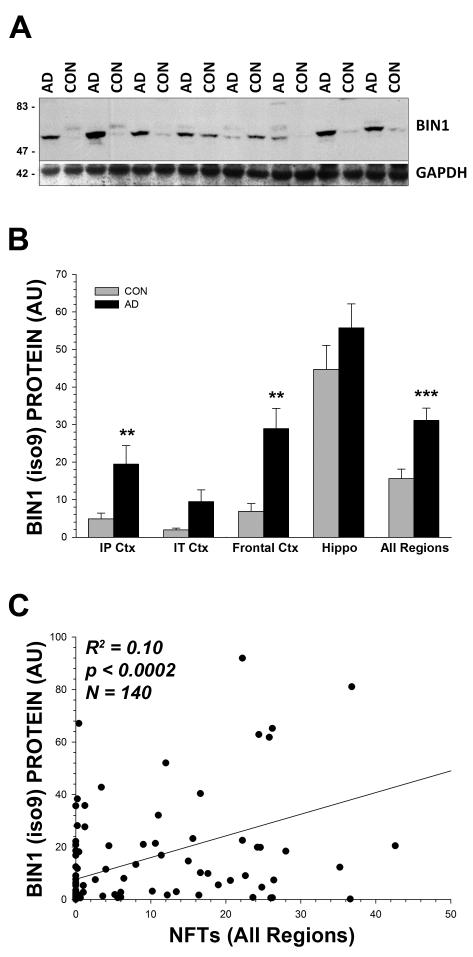
Recent genome wide association studies have implicated bridging integrator 1 (BIN1) as a late-onset Alzheimer's disease (AD) susceptibility gene. There are at least 15 different known isoforms of BIN1, with many being expressed in the brain including the longest isoform (iso1), which is brain-specific and localizes to axon initial segments and nodes of Ranvier. It is currently unknown what role BIN1 plays in AD. We analyzed BIN1 protein expression from a large number (n = 71) of AD cases and controls from five different brain regions (hippocampus, inferior parietal cortex, inferior temporal cortex, frontal cortex (BA9), and superior and middle temporal gyri). We found that the amount of the largest isoform of BIN1 was significantly reduced in the AD brain compared to age-matched controls, and smaller BIN1 isoforms were significantly increased. Further, BIN1 was significantly correlated with the amount of neurofibrillary tangle (NFT) pathology but not with either diffuse or neuritic plaques, or with the amount of amyloid-β peptide. BIN1 is known to be abnormally expressed in another human disease, myotonic dystrophy, which also features prominent NFT pathology. These data suggest that BIN1 is likely involved in AD as a modulator of NFT pathology, and that this role may extend to other human diseases that feature tau pathology.
Keywords: Alzheimer's disease; ZNF9; amyloid-β peptide; cellular nucleic acid binding protein; myotonic dystrophy; tau.
Figures

Similar articles
-
Aberrant accrual of BIN1 near Alzheimer's disease amyloid deposits in transgenic models.Brain Pathol. 2019 Jul;29(4):485-501. doi: 10.1111/bpa.12687. Epub 2018 Dec 27. Brain Pathol. 2019. PMID: 30506549 Free PMC article.
-
The Mechanistic Role of Bridging Integrator 1 (BIN1) in Alzheimer's Disease.Cell Mol Neurobiol. 2021 Oct;41(7):1431-1440. doi: 10.1007/s10571-020-00926-y. Epub 2020 Jul 27. Cell Mol Neurobiol. 2021. PMID: 32719966 Free PMC article. Review.
-
Intracellular clusterin interacts with brain isoforms of the bridging integrator 1 and with the microtubule-associated protein Tau in Alzheimer's disease.PLoS One. 2014 Jul 22;9(7):e103187. doi: 10.1371/journal.pone.0103187. eCollection 2014. PLoS One. 2014. PMID: 25051234 Free PMC article.
-
BIN1 protein isoforms are differentially expressed in astrocytes, neurons, and microglia: neuronal and astrocyte BIN1 are implicated in tau pathology.Mol Neurodegener. 2020 Jul 29;15(1):44. doi: 10.1186/s13024-020-00387-3. Mol Neurodegener. 2020. PMID: 32727516 Free PMC article.
-
BIN1 in the Pursuit of Ousting the Alzheimer's Reign: Impact on Amyloid and Tau Neuropathology.Neurotox Res. 2023 Dec;41(6):698-707. doi: 10.1007/s12640-023-00670-3. Epub 2023 Oct 17. Neurotox Res. 2023. PMID: 37847429 Review.
Cited by
-
Early Life Stress and Epigenetics in Late-onset Alzheimer's Dementia: A Systematic Review.Curr Genomics. 2018 Nov;19(7):522-602. doi: 10.2174/1389202919666171229145156. Curr Genomics. 2018. PMID: 30386171 Free PMC article. Review.
-
Aberrant accrual of BIN1 near Alzheimer's disease amyloid deposits in transgenic models.Brain Pathol. 2019 Jul;29(4):485-501. doi: 10.1111/bpa.12687. Epub 2018 Dec 27. Brain Pathol. 2019. PMID: 30506549 Free PMC article.
-
Comprehensive Review on Alzheimer's Disease: Causes and Treatment.Molecules. 2020 Dec 8;25(24):5789. doi: 10.3390/molecules25245789. Molecules. 2020. PMID: 33302541 Free PMC article. Review.
-
The Mechanistic Role of Bridging Integrator 1 (BIN1) in Alzheimer's Disease.Cell Mol Neurobiol. 2021 Oct;41(7):1431-1440. doi: 10.1007/s10571-020-00926-y. Epub 2020 Jul 27. Cell Mol Neurobiol. 2021. PMID: 32719966 Free PMC article. Review.
-
Upregulation of RIN3 induces endosomal dysfunction in Alzheimer's disease.Transl Neurodegener. 2020 Jun 18;9(1):26. doi: 10.1186/s40035-020-00206-1. Transl Neurodegener. 2020. PMID: 32552912 Free PMC article.
References
-
- Harold D, Abraham R, Hollingworth P, Sims R, Gerrish A, Hamshere ML, Pahwa JS, Moskvina V, Dowzell K, Williams A, Jones N, Thomas C, Stretton A, Morgan AR, Lovestone S, Powell J, Proitsi P, Lupton MK, Brayne C, Rubinsztein DC, Gill M, Lawlor B, Lynch A, Morgan K, Brown KS, Passmore PA, Craig D, McGuinness B, Todd S, Holmes C, Mann D, Smith AD, Love S, Kehoe PG, Hardy J, Mead S, Fox N, Rossor M, Collinge J, Maier W, Jessen F, Schurmann B, van den Bussche H, Heuser I, Kornhuber J, Wiltfang J, Dichgans M, Frolich L, Hampel H, Hull M, Rujescu D, Goate AM, Kauwe JS, Cruchaga C, Nowotny P, Morris JC, Mayo K, Sleegers K, Bettens K, Engelborghs S, De Deyn PP, Van Broeckhoven C, Livingston G, Bass NJ, Gurling H, McQuillin A, Gwilliam R, Deloukas P, Al-Chalabi A, Shaw CE, Tsolaki M, Singleton AB, Guerreiro R, Muhleisen TW, Nothen MM, Moebus S, Jockel KH, Klopp N, Wichmann HE, Carrasquillo MM, Pankratz VS, Younkin SG, Holmans PA, O’Donovan M, Owen MJ, Williams J. Genome-wide association study identifies variants at CLU and PICALM associated with Alzheimer’s disease. Nat Genet. 2009;41:1088–1093. - PMC - PubMed
-
- Lambert JC, Heath S, Even G, Campion D, Sleegers K, Hiltunen M, Combarros O, Zelenika D, Bullido MJ, Tavernier B, Letenneur L, Bettens K, Berr C, Pasquier F, Fievet N, Barberger-Gateau P, Engelborghs S, De Deyn P, Mateo I, Franck A, Helisalmi S, Porcellini E, Hanon O, de Pancorbo MM, Lendon C, Dufouil C, Jaillard C, Leveillard T, Alvarez V, Bosco P, Mancuso M, Panza F, Nacmias B, Bossu P, Piccardi P, Annoni G, Seripa D, Galimberti D, Hannequin D, Licastro F, Soininen H, Ritchie K, Blanche H, Dartigues JF, Tzourio C, Gut I, Van Broeckhoven C, Alperovitch A, Lathrop M, Amouyel P. Genome-wide association study identifies variants at CLU and CR1 associated with Alzheimer’s disease. Nat Genet. 2009;41:1094–1099. - PubMed
-
- Seshadri S, Fitzpatrick AL, Ikram MA, DeStefano AL, Gudnason V, Boada M, Bis JC, Smith AV, Carassquillo MM, Lambert JC, Harold D, Schrijvers EM, Ramirez-Lorca R, Debette S, Longstreth WT, Jr., Janssens AC, Pankratz VS, Dartigues JF, Hollingworth P, Aspelund T, Hernandez I, Beiser A, Kuller LH, Koudstaal PJ, Dickson DW, Tzourio C, Abraham R, Antunez C, Du Y, Rotter JI, Aulchenko YS, Harris TB, Petersen RC, Berr C, Owen MJ, Lopez-Arrieta J, Varadarajan BN, Becker JT, Rivadeneira F, Nalls MA, Graff-Radford NR, Campion D, Auerbach S, Rice K, Hofman A, Jonsson PV, Schmidt H, Lathrop M, Mosley TH, Au R, Psaty BM, Uitterlinden AG, Farrer LA, Lumley T, Ruiz A, Williams J, Amouyel P, Younkin SG, Wolf PA, Launer LJ, Lopez OL, van Duijn CM, Breteler MM. Genome-wide analysis of genetic loci associated with Alzheimer disease. JAMA: the journal of the American Medical Association. 2010;303:1832–1840. - PMC - PubMed
-
- Sakamuro D, Elliott KJ, Wechsler-Reya R, Prendergast GC. BIN1 is a novel MYC-interacting protein with features of a tumour suppressor. Nat Genet. 1996;14:69–77. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- UL1 TR000117/TR/NCATS NIH HHS/United States
- N01-HD-9-0011/HD/NICHD NIH HHS/United States
- AG028383/AG/NIA NIH HHS/United States
- NS083692/NS/NINDS NIH HHS/United States
- AG005119/AG/NIA NIH HHS/United States
- P50 AG016573/AG/NIA NIH HHS/United States
- R21 NS083692/NS/NINDS NIH HHS/United States
- P50 AG025688/AG/NIA NIH HHS/United States
- P30 AG028383/AG/NIA NIH HHS/United States
- GM103486/GM/NIGMS NIH HHS/United States
- P20 GM103486/GM/NIGMS NIH HHS/United States
- NS058382/NS/NINDS NIH HHS/United States
- P01 AG005119/AG/NIA NIH HHS/United States
- TL1 TR000115/TR/NCATS NIH HHS/United States
- R01 NS058382/NS/NINDS NIH HHS/United States
- T32 NS007480/NS/NINDS NIH HHS/United States
- P50AG16573/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
