

The NB-LRR proteins RGA4 and RGA5 interact functionally and physically to confer disease resistance
- PMID: 25024433
- PMCID: PMC4195788
- DOI: 10.15252/embj.201487923
The NB-LRR proteins RGA4 and RGA5 interact functionally and physically to confer disease resistance
Abstract
Plant resistance proteins of the class of nucleotide-binding and leucine-rich repeat domain proteins (NB-LRRs) are immune sensors which recognize pathogen-derived molecules termed avirulence (AVR) proteins. We show that RGA4 and RGA5, two NB-LRRs from rice, interact functionally and physically to mediate resistance to the fungal pathogen Magnaporthe oryzae and accomplish different functions in AVR recognition. RGA4 triggers an AVR-independent cell death that is repressed in the presence of RGA5 in both rice protoplasts and Nicotiana benthamiana. Upon recognition of the pathogen effector AVR-Pia by direct binding to RGA5, repression is relieved and cell death occurs. RGA4 and RGA5 form homo- and hetero-complexes and interact through their coiled-coil domains. Localization studies in rice protoplast suggest that RGA4 and RGA5 localize to the cytosol. Upon recognition of AVR-Pia, neither RGA4 nor RGA5 is re-localized to the nucleus. These results establish a model for the interaction of hetero-pairs of NB-LRRs in plants: RGA4 mediates cell death activation, while RGA5 acts as a repressor of RGA4 and as an AVR receptor.
Keywords: Magnaporthe oryzae; pathogen recognition; plant immunity; resistance protein; rice.
© 2014 The Authors.
Figures
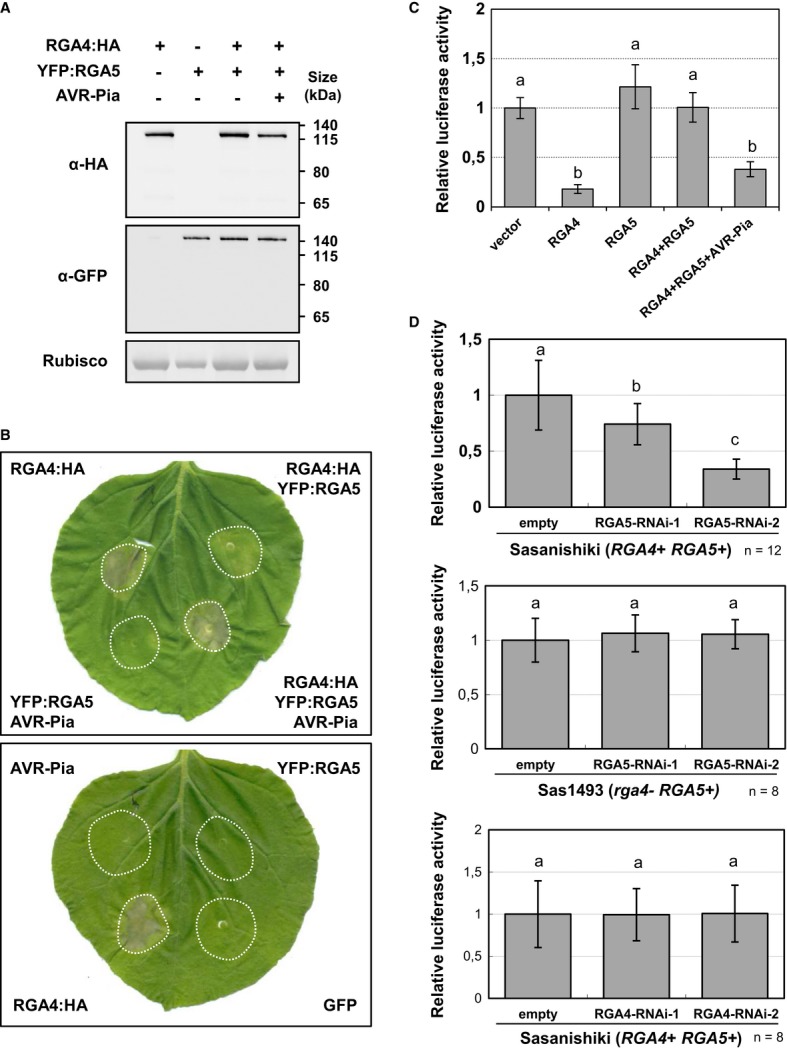
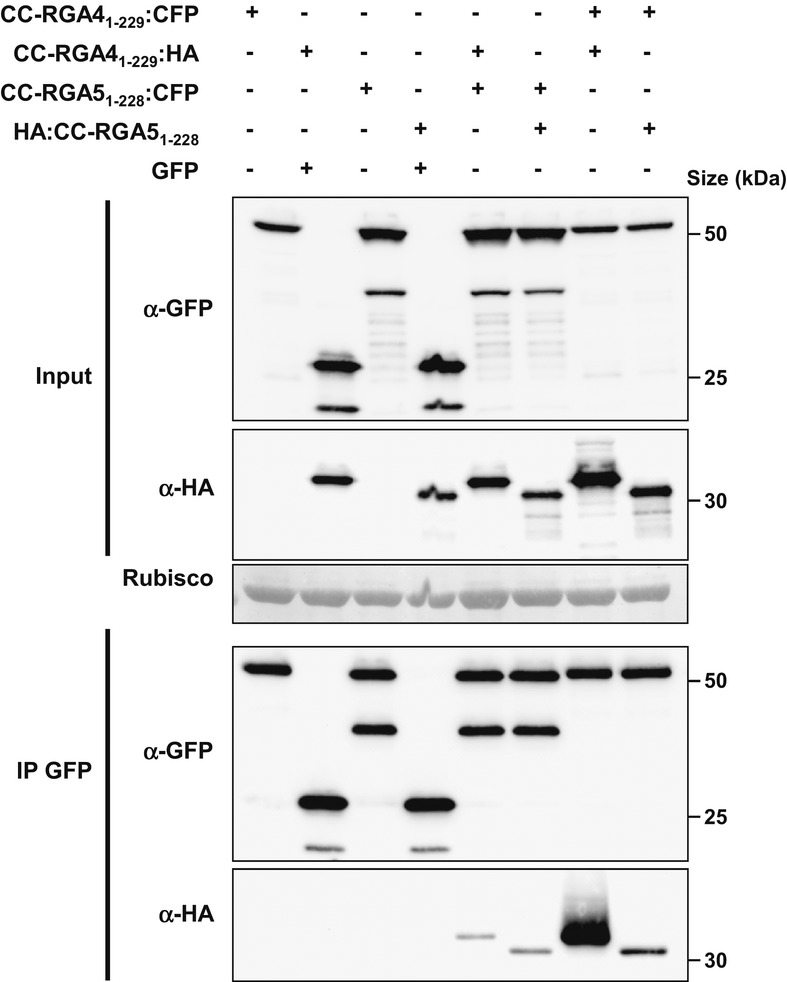
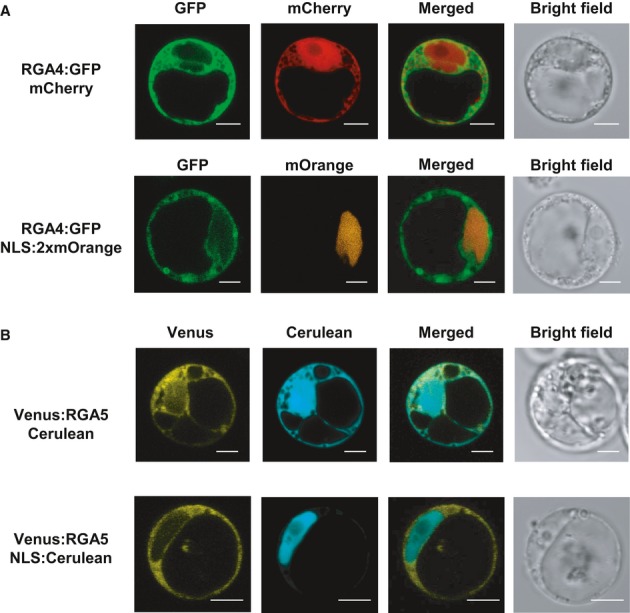
RGA4:HA, YFP:RGA5, and AVR-Pia constructs were transiently expressed in Nicotiana benthamiana leaves by A. tumefaciens infiltration. Proteins were extracted 24 h after infiltration, and protein extracts were analyzed by immunoblotting with anti-GFP (α-GFP) and anti-HA (α-HA) antibodies. Ponceau staining of Rubisco small subunit was used to verify equal protein loading.
The indicated combinations of fusion proteins were transiently expressed in N. benthamiana. The RGA4:HA strain was infiltrated at OD600 of 0.2, while strains carrying all other constructs were infiltrated at OD600 of 0.4. A bacterial strain carrying the GFP construct was used to equilibrate the total OD600 to 1 before infiltration. Cell death induced by the different combinations was visualized 3 days after infiltration. Equivalent results were obtained in at least three independent experiments.
Leaf protoplasts of the susceptible rice cultivar Himenomochi (pia) were transfected with a plasmid for constitutive LUC expression in combination with constructs allowing expression of RGA4, RGA5, RGA4+RGA5, or RGA4+RGA5+AVR-Pia. LUC activity was determined in protein extracts of protoplast samples harvested 40 h after transfection. Average values and standard deviations were calculated from three replicate samples, and values were normalized with respect to the average values of the empty vector samples. The experiment was repeated three times with equivalent results. Different letters indicate groups that are significantly different (P < 0.05 in pairwise t-test).
Rice protoplast of the resistant rice variety Sasanishiki or of the rga4 mutant line Sas1493 was co-transfected with RNAi constructs directed against RGA4 or RGA5 and the luciferase reporter gene. Luc activity was determined and analyzed as in (C). Average values and standard deviations were calculated from the indicated number of replicate samples (n). The experiment was repeated three times with equivalent results.
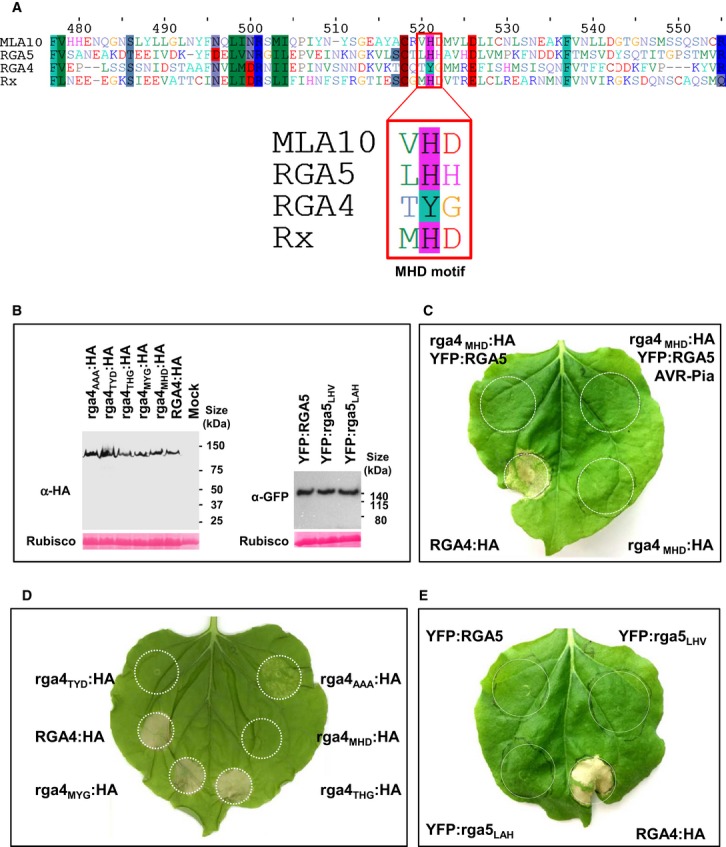
Alignment of a portion of the ARC2 domain sequences of RGA4, RGA5, Rx, and MLA10 containing the MHD motif (red box).
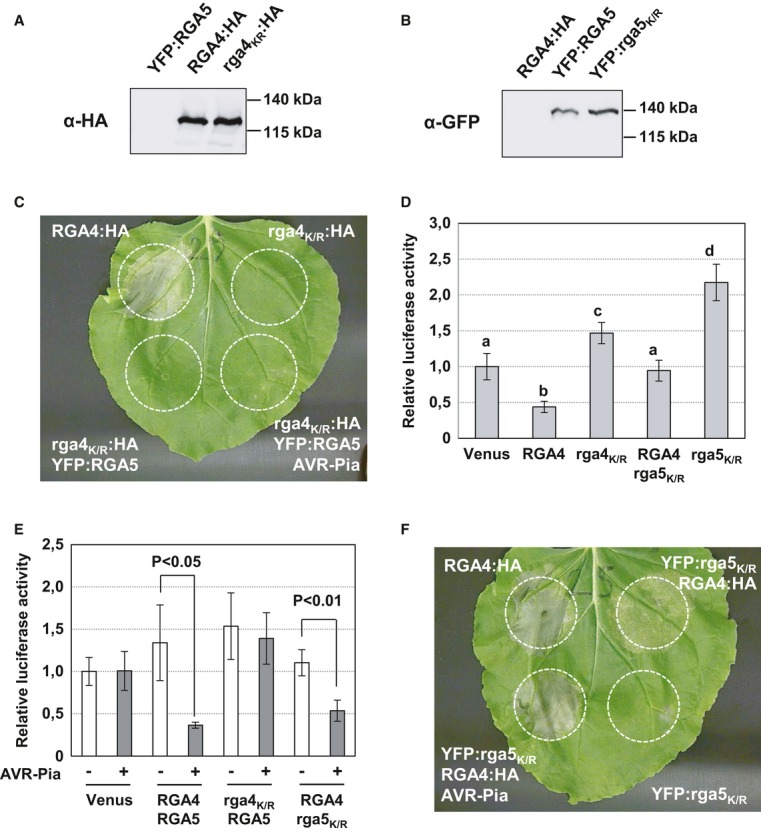
Proteins were extracted 24 h after infiltration of the indicated RGA4:HA or YFP:RGA5 constructs in N. benthamiana and analyzed by immunoblotting with anti-HA or anti-GFP antibodies (α-HA and α-GFP). Ponceau staining of Rubisco small subunit was used to verify equal protein loading.
The rga4MHD:HA strain harboring a construct for expression of an RGA4 variant with the MHD consensus motif was infiltrated alone and in combination with YFP:RGA5 or YFP:RGA5 and AVR-Pia strains in N. benthamiana. Cell death was visualized 3 days after infiltration. The same results were obtained in at least three independent experiments.
The RGA4 MHD mutant constructs rga4MHD:HA, rga4MYG:HA, rga4THG:HA, rga4TYD:HA and rga4AAA:HA were expressed in N. benthamiana and analyzed for cell death induction.
YFP:RGA5, YFP:rga5LHV, YFP:rga5LAH, and RGA4:HA were expressed in N. benthamiana, and cell death was visualized 3 days after infiltration. Similar results were obtained in at least three independent experiments.
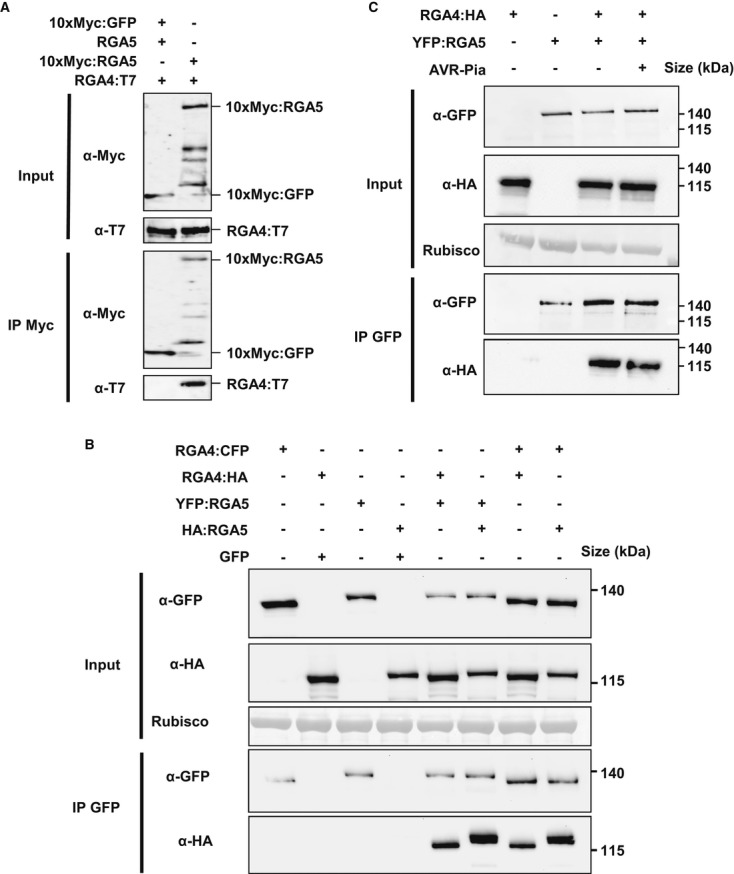
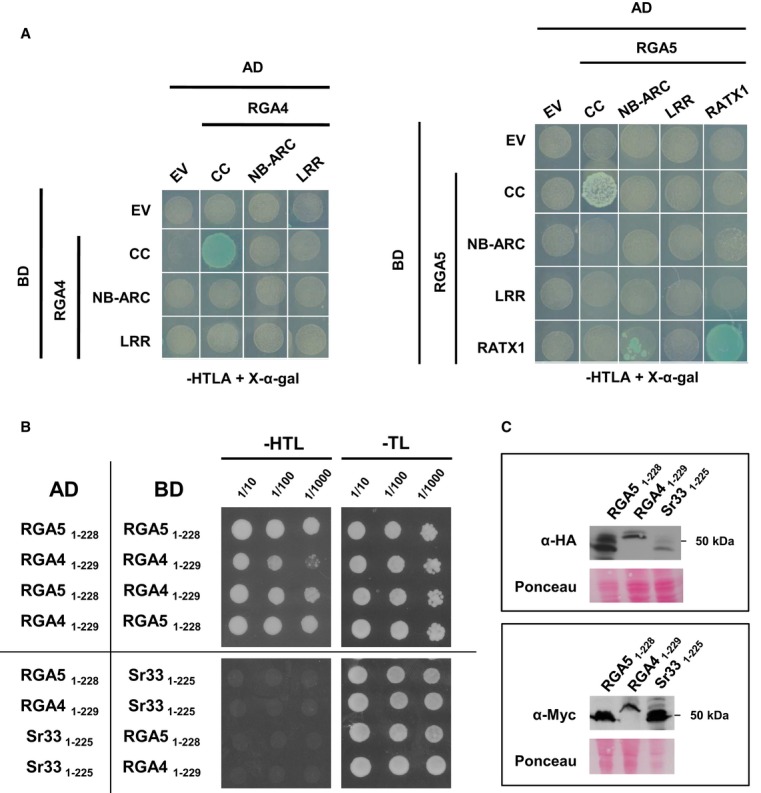
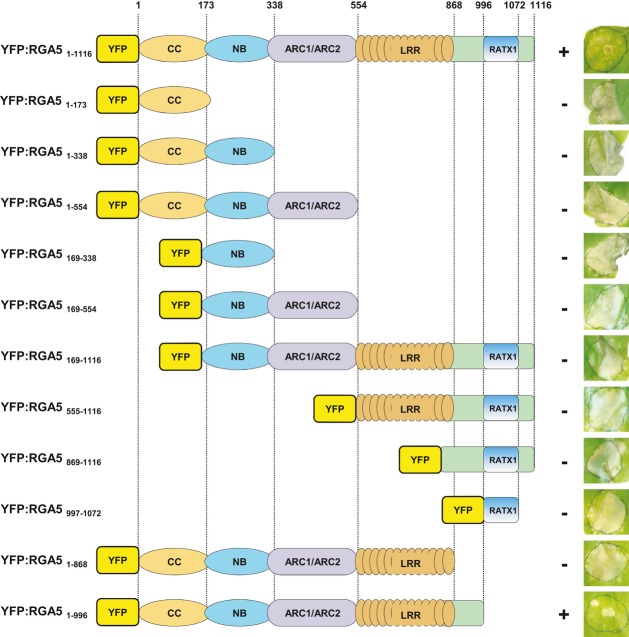
Pairwise interaction of CC, NB-ARC, and LRR domains from RGA4 and RGA5, as well as the RGA5 RATX1 domain (see Supplementary Fig S9A for domain borders) was evaluated by blue coloration and growth of yeast cells on basal media lacking Trp, Leu, Ade, and His and supplemented with 5-bromo-4-chloro-3-indolyl α-D-galactopyranoside (X-a-gal). Pictures were taken after 3 days of growth.
RGA4 and RGA5 CC domain homo- and hetero-complexes. Dilution series of yeast cells expressing GAL4-AD and GAL4-BD fusions of RGA5 1–228 and/or RGA4 1–229 CC domains on non-selective synthetic media lacking Trp and Leu (-TL) and selective media additionally lacking His (-HTL). The wheat Sr33 (1–225) CC domain was used as a negative control. Pictures were taken after 3 days of growth. Growth on -HTL media is indicative of interaction.
GAL4-AD and -BD fusions of CC domains of RGA4, RGA5, and Sr33 were detected by immunoblotting of yeast protein extracts with, respectively, anti-HA and anti-Myc antibodies. Equal loading was verified by red Ponceau staining.
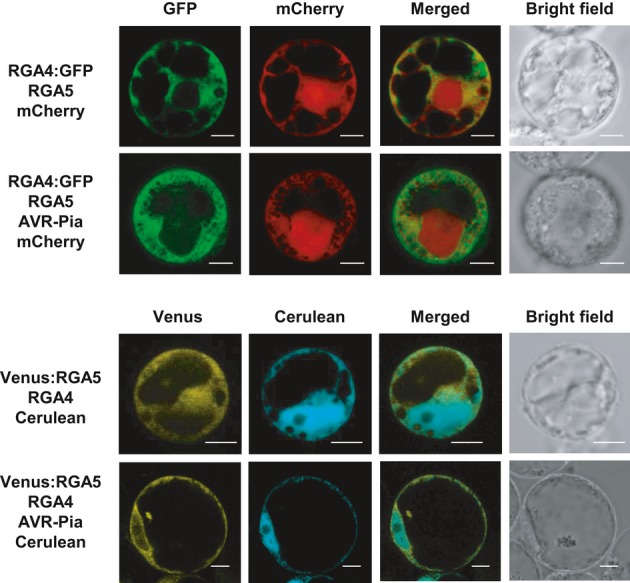
References
-
- Akamatsu A, Wong HL, Fujiwara M, Okuda J, Nishide K, Uno K, Imai K, Umemura K, Kawasaki T, Kawano Y, Shimamoto K. An OsCEBiP/OsCERK1-OsRacGEF1-OsRac1 module is an essential early component of chitin-induced rice immunity. Cell Host Microbe. 2013;13:465–476. - PubMed
-
- Ausubel FM. Are innate immune signaling pathways in plants and animals conserved? Nat Immunol. 2005;6:973–979. - PubMed
-
- Baba A, Hasezawa S, Syōno K. Cultivation of rice protoplasts and their transformation mediated by agrobacterium spheroplasts. Plant Cell Physiol. 1986;27:463–471.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
