

Investigating the structural impact of the glutamine repeat in huntingtin assembly
- PMID: 25028883
- PMCID: PMC4104047
- DOI: 10.1016/j.bpj.2014.06.002
Investigating the structural impact of the glutamine repeat in huntingtin assembly
Abstract
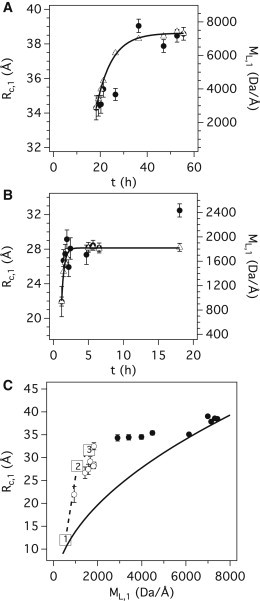
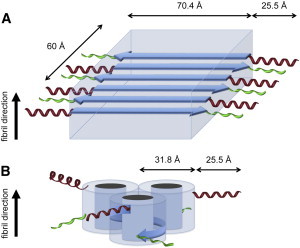
Acquiring detailed structural information about the various aggregation states of the huntingtin-exon1 protein (Htt-exon1) is crucial not only for identifying the true nature of the neurotoxic species responsible for Huntington's disease (HD) but also for designing effective therapeutics. Using time-resolved small-angle neutron scattering (TR-SANS), we followed the conformational changes that occurred during fibrillization of the pathologic form of Htt-exon1 (NtQ42P10) and compared the results with those obtained for the wild-type (NtQ22P10). Our results show that the aggregation pathway of NtQ22P10 is very different from that of NtQ42P10, as the initial steps require a monomer to 7-mer transition stage. In contrast, the earliest species identified for NtQ42P10 are monomer and dimer. The divergent pathways ultimately result in NtQ22P10 fibrils that possess a packing arrangement consistent with the common amyloid sterical zipper model, whereas NtQ42P10 fibrils present a better fit to the Perutz β-helix structural model. The structural details obtained by TR-SANS should help to delineate the key mechanisms that underpin Htt-exon1 aggregation leading to HD.
Copyright © 2014 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures





References
-
- MacDonald M.E., Ambrose C.M., Duyao M.P., Myers R.H., Lin C., The Huntington’s Disease Collaborative Research Group A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington’s disease chromosomes. Cell. 1993;72:971–983. - PubMed
-
- Sawa A., Nagata E., Snyder S.H. Huntingtin is cleaved by caspases in the cytoplasm and translocated to the nucleus via perinuclear sites in Huntington’s disease patient lymphoblasts. Neurobiol. Dis. 2005;20:267–274. - PubMed
-
- Graham R.K., Deng Y., Hayden M.R. Cleavage at the caspase-6 site is required for neuronal dysfunction and degeneration due to mutant huntingtin. Cell. 2006;125:1179–1191. - PubMed
-
- Darnell G., Orgel J.P.R.O., Meredith S.C. Flanking polyproline sequences inhibit β-sheet structure in polyglutamine segments by inducing PPII-like helix structure. J. Mol. Biol. 2007;374:688–704. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
