

The cancer glycocalyx mechanically primes integrin-mediated growth and survival
- PMID: 25030168
- PMCID: PMC4487551
- DOI: 10.1038/nature13535
The cancer glycocalyx mechanically primes integrin-mediated growth and survival
Abstract
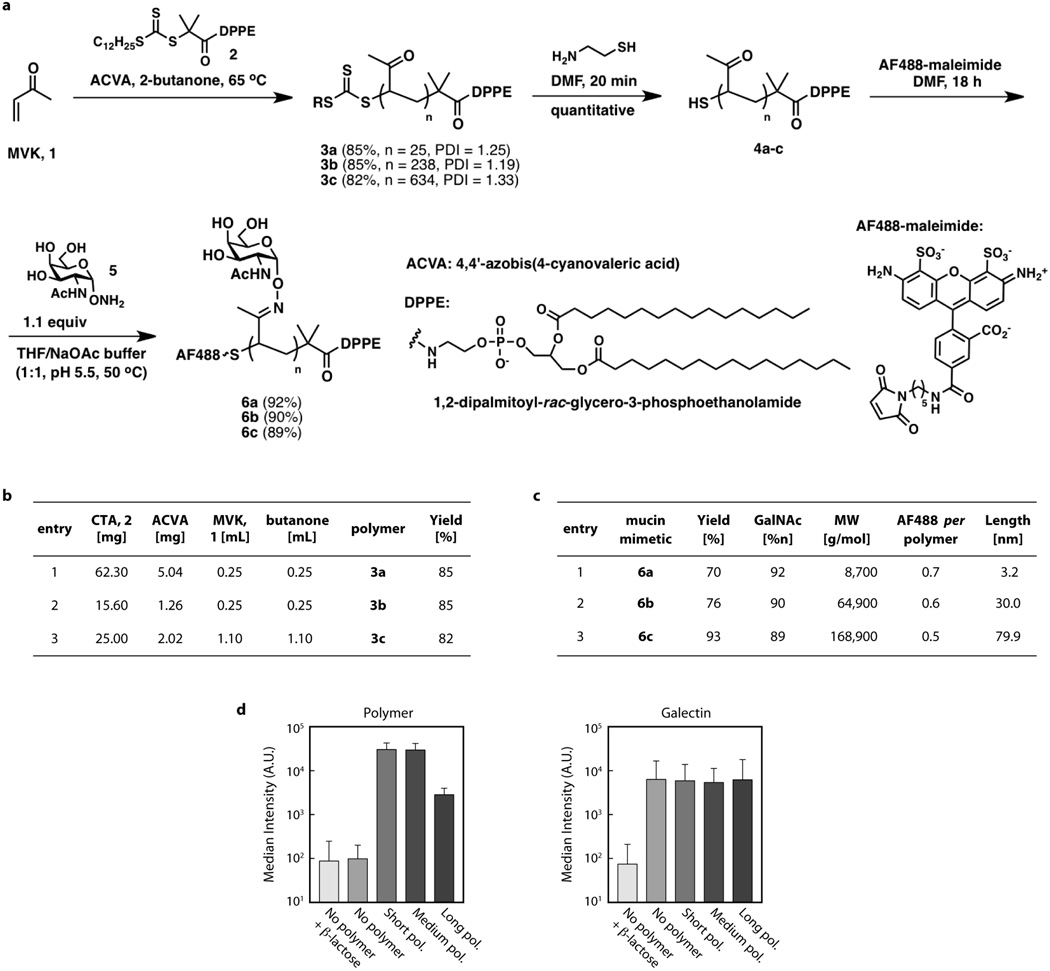
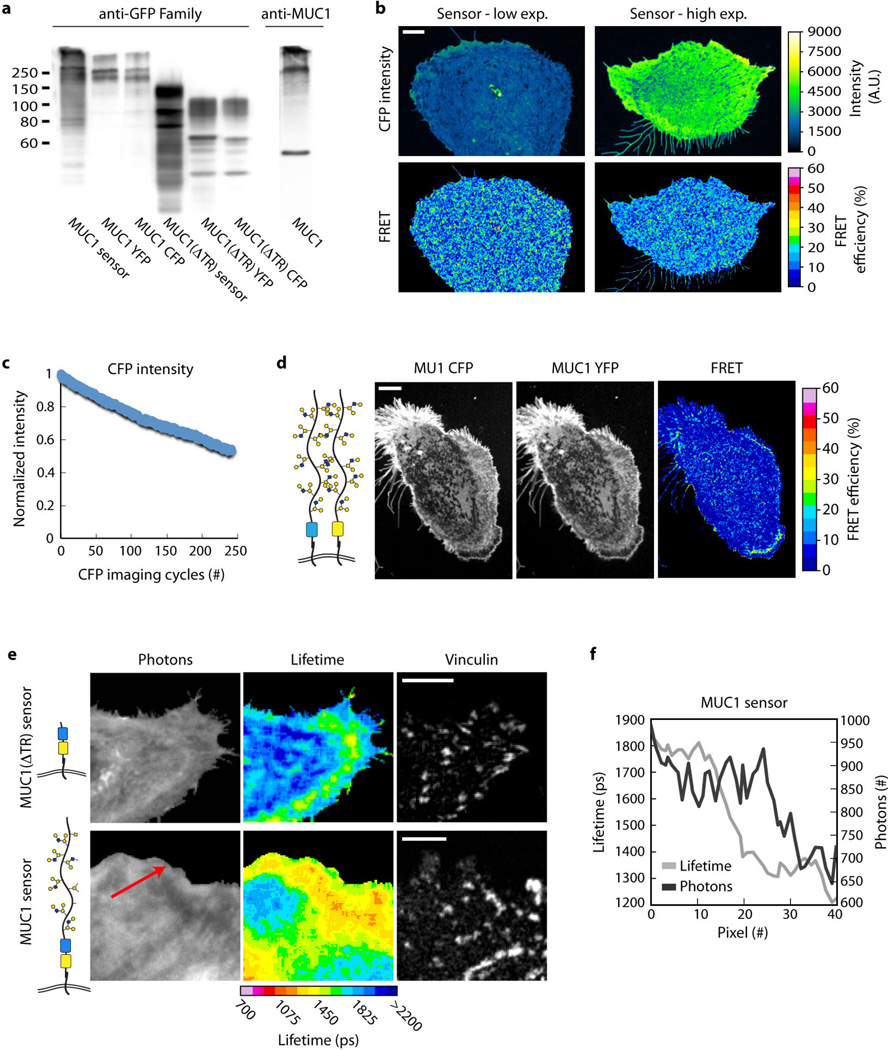
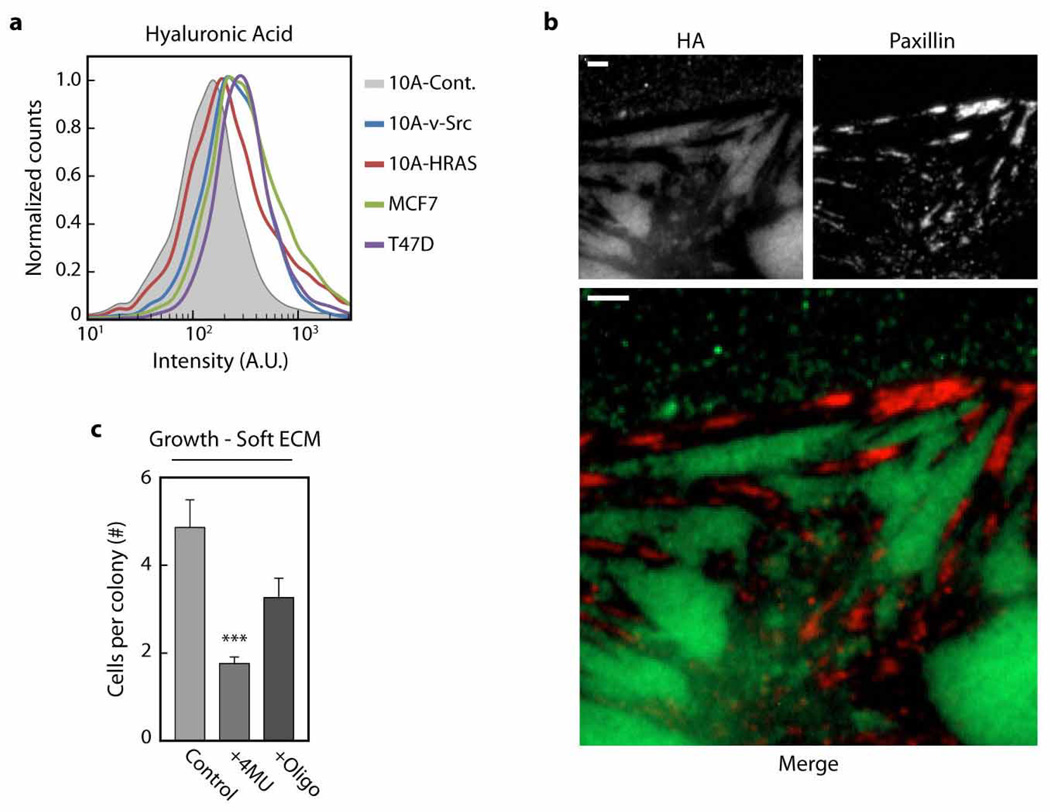
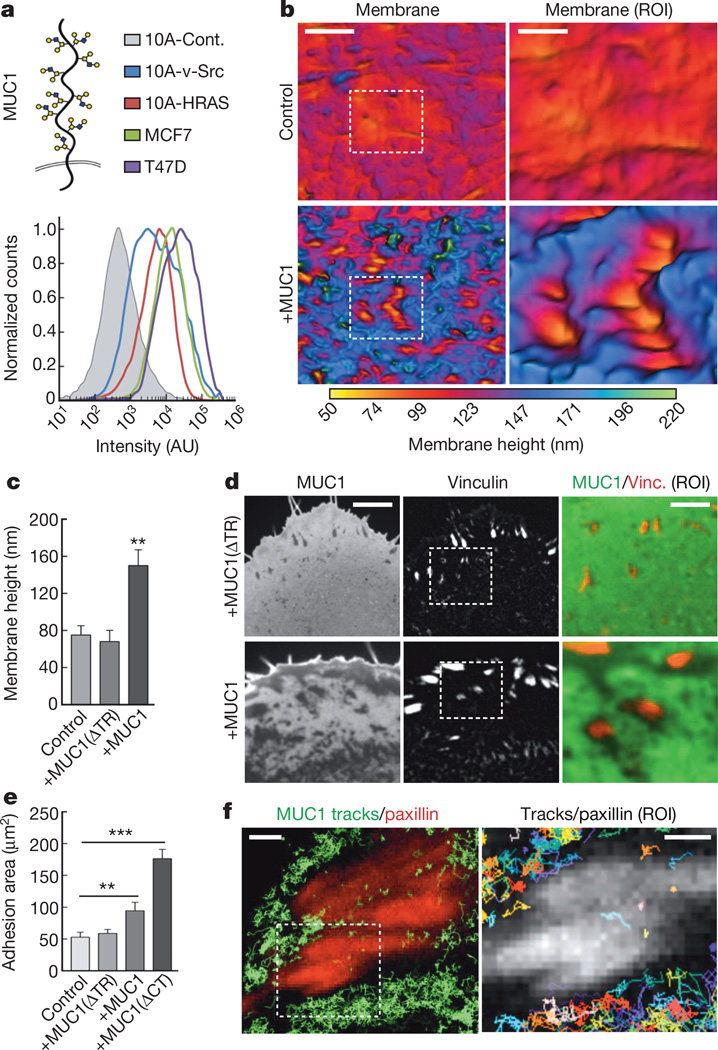
Malignancy is associated with altered expression of glycans and glycoproteins that contribute to the cellular glycocalyx. We constructed a glycoprotein expression signature, which revealed that metastatic tumours upregulate expression of bulky glycoproteins. A computational model predicted that these glycoproteins would influence transmembrane receptor spatial organization and function. We tested this prediction by investigating whether bulky glycoproteins in the glycocalyx promote a tumour phenotype in human cells by increasing integrin adhesion and signalling. Our data revealed that a bulky glycocalyx facilitates integrin clustering by funnelling active integrins into adhesions and altering integrin state by applying tension to matrix-bound integrins, independent of actomyosin contractility. Expression of large tumour-associated glycoproteins in non-transformed mammary cells promoted focal adhesion assembly and facilitated integrin-dependent growth factor signalling to support cell growth and survival. Clinical studies revealed that large glycoproteins are abundantly expressed on circulating tumour cells from patients with advanced disease. Thus, a bulky glycocalyx is a feature of tumour cells that could foster metastasis by mechanically enhancing cell-surface receptor function.
Figures











Comment in
-
Cancer: Sugar-coated cell signalling.Nature. 2014 Jul 17;511(7509):298-9. doi: 10.1038/nature13506. Epub 2014 Jun 25. Nature. 2014. PMID: 25030162 No abstract available.
References
-
- Haltiwanger RS, Lowe JB. Role of glycosylation in development. Annu. Rev. Biochem. 2004;73:491–537. - PubMed
-
- Varki A, Kannagi R, Toole BP. In: Essentials Glycobiol. Varki A, et al., editors. Cold Spring Harbor Laboratory Press; 2009. http://www.ncbi.nlm.nih.gov/books/NBK1963/
-
- Walker JL, Fournier AK, Assoian RK. Regulation of growth factor signaling and cell cycle progression by cell adhesion and adhesion-dependent changes in cellular tension. Cytokine Growth Factor Rev. 2005;16:395–405. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- U54CA163155-01/CA/NCI NIH HHS/United States
- K99 EB013446-02/EB/NIBIB NIH HHS/United States
- R01 CA138818/CA/NCI NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- U54CA143836-01/CA/NCI NIH HHS/United States
- R00 EB013446/EB/NIBIB NIH HHS/United States
- GM59907/GM/NIGMS NIH HHS/United States
- K99 EB013446/EB/NIBIB NIH HHS/United States
- U54 CA163155/CA/NCI NIH HHS/United States
- R01 GM059907/GM/NIGMS NIH HHS/United States
- U01 ES019458/ES/NIEHS NIH HHS/United States
- U01 CA151925/CA/NCI NIH HHS/United States
- S10 RR026758/RR/NCRR NIH HHS/United States
- T32 GM066698/GM/NIGMS NIH HHS/United States
- R01 AI082292/AI/NIAID NIH HHS/United States
- U54 CA143836/CA/NCI NIH HHS/United States
- 2R01GM059907-13/GM/NIGMS NIH HHS/United States
- 1U01 ES019458-01/ES/NIEHS NIH HHS/United States
- CA138818-01A1/CA/NCI NIH HHS/United States
- AI082292-03A1/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
