

Hypoactivation of CRF receptors, predominantly type 2, in the medial-posterior BNST is vital for adequate maternal behavior in lactating rats
- PMID: 25031406
- PMCID: PMC4099544
- DOI: 10.1523/JNEUROSCI.4220-13.2014
Hypoactivation of CRF receptors, predominantly type 2, in the medial-posterior BNST is vital for adequate maternal behavior in lactating rats
Abstract
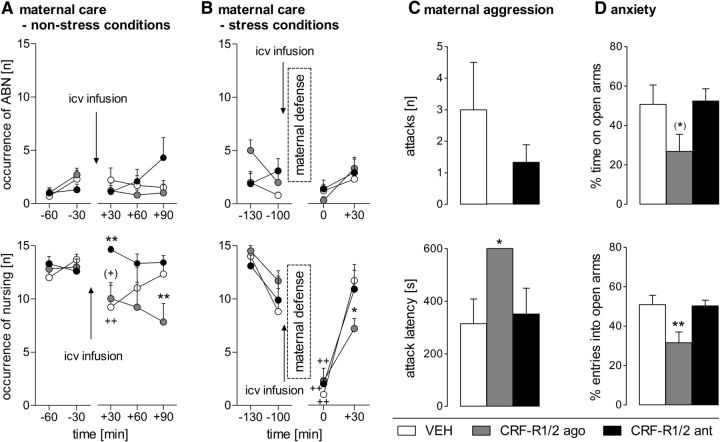
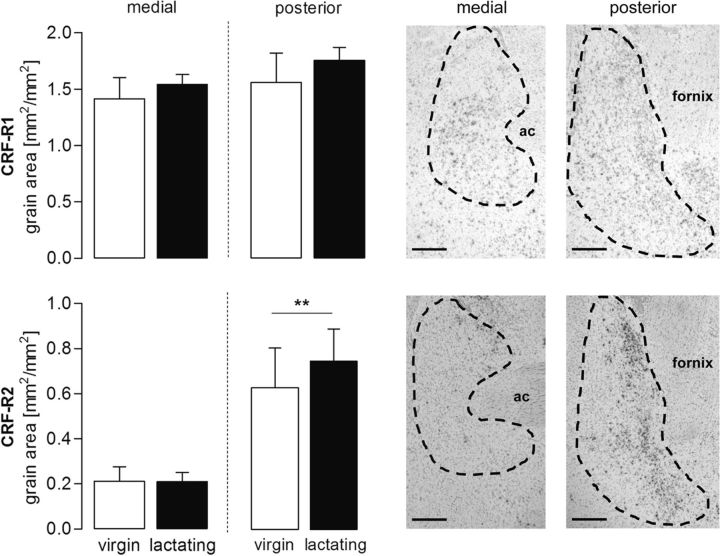
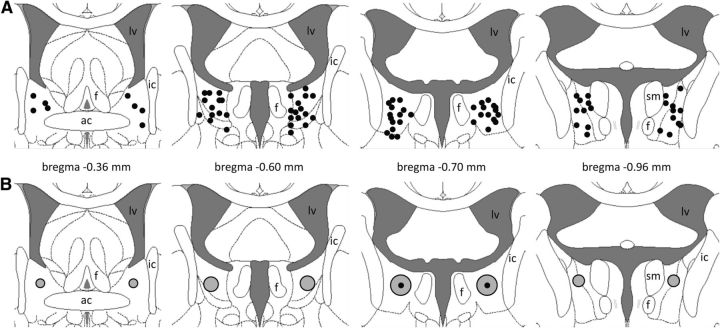
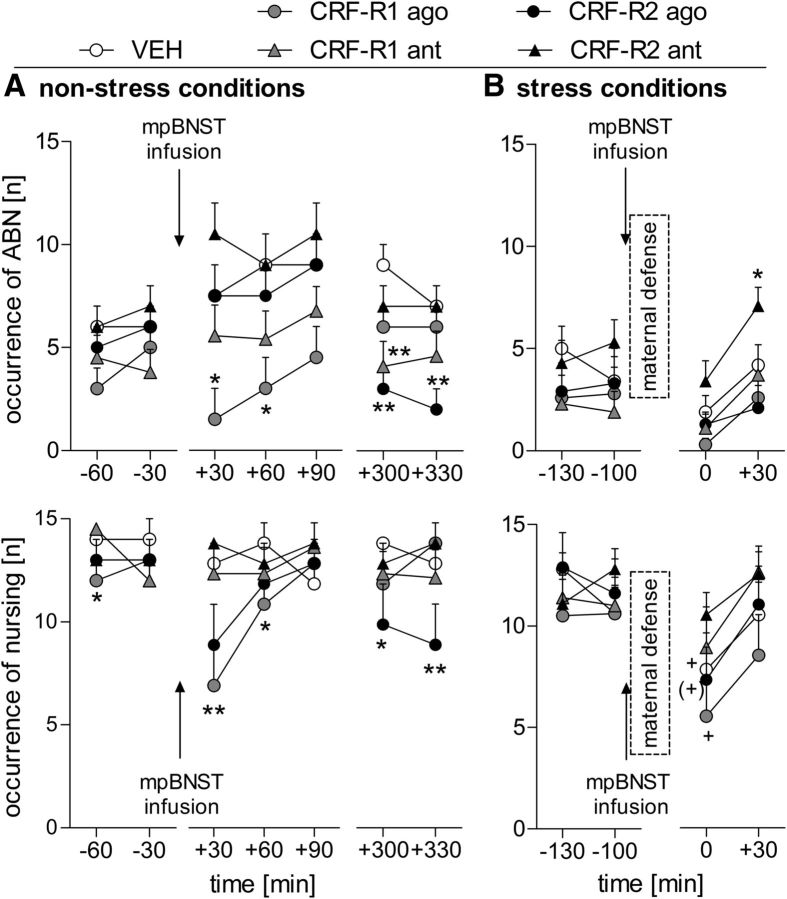
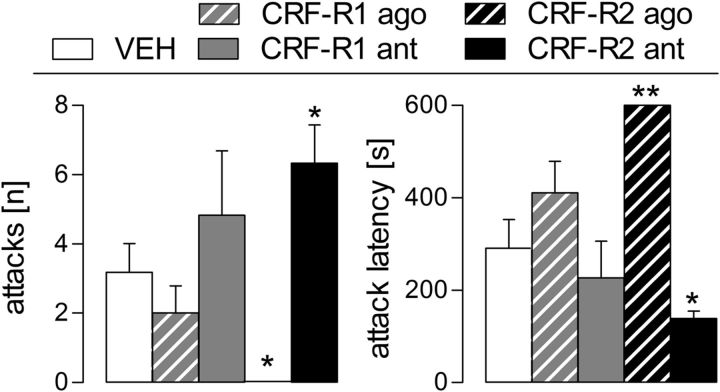
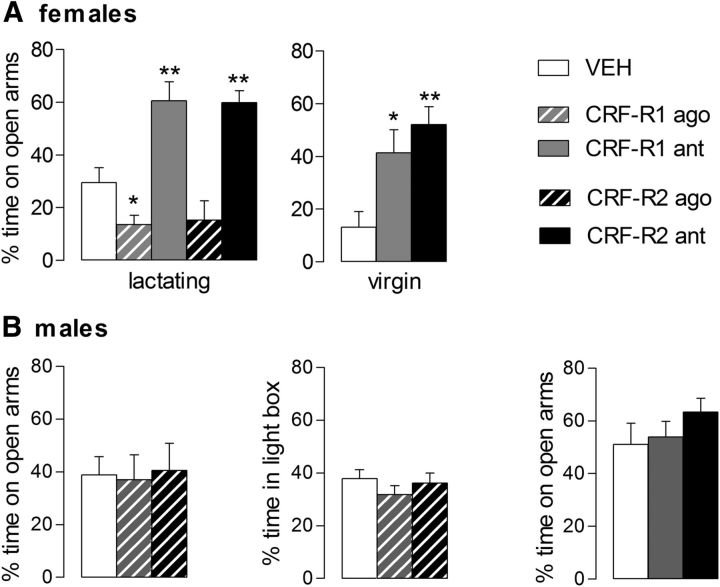
Maternal behavior ensures the proper development of the offspring. In lactating mammals, maternal behavior is impaired by stress, the physiological consequence of central corticotropin-releasing factor receptor (CRF-R) activation. However, which CRF-R subtype in which specific brain area(s) mediates this effect is unknown. Here we confirmed that an intracerebroventricularly injected nonselective CRF-R antagonist enhances, whereas an agonist impairs, maternal care. The agonist also prolonged the stress-induced decrease in nursing, reduced maternal aggression and increased anxiety-related behavior. Focusing on the bed nucleus of the stria terminalis (BNST), CRF-R1 and CRF-R2 mRNA expression did not differ in virgin versus lactating rats. However, CRF-R2 mRNA was more abundant in the posterior than in the medial BNST. Pharmacological manipulations within the medial-posterior BNST showed that both CRF-R1 and CRF-R2 agonists reduced arched back nursing (ABN) rapidly and after a delay, respectively. After stress, both antagonists prevented the stress-induced decrease in nursing, with the CRF-R2 antagonist actually increasing ABN. During the maternal defense test, maternal aggression was abolished by the CRF-R2, but not the CRF-R1, agonist. Anxiety-related behavior was increased by the CRF-R1 agonist and reduced by both antagonists. Both antagonists were also effective in virgin females but not in males, revealing a sexual dimorphism in the regulation of anxiety within the medial-posterior BNST. In conclusion, the detrimental effects of increased CRF-R activation on maternal behavior are mediated via CRF-R2 and, to a lesser extent, via CRF-R1 in the medial-posterior BNST in lactating rats. Moreover, both CRF-R1 and CRF-R2 regulate anxiety in females independently of their reproductive status.
Copyright © 2014 Klampfl et al.
Figures
Similar articles
-
CRF-R1 activation in the anterior-dorsal BNST induces maternal neglect in lactating rats via an HPA axis-independent central mechanism.Psychoneuroendocrinology. 2016 Feb;64:89-98. doi: 10.1016/j.psyneuen.2015.11.015. Epub 2015 Nov 26. Psychoneuroendocrinology. 2016. PMID: 26630389 Free PMC article.
-
Brain CRF-binding protein modulates aspects of maternal behavior under stressful conditions and supports a hypo-anxious state in lactating rats.Horm Behav. 2016 Aug;84:136-44. doi: 10.1016/j.yhbeh.2016.06.009. Epub 2016 Jun 28. Horm Behav. 2016. PMID: 27368148
-
Maternal stress and the MPOA: Activation of CRF receptor 1 impairs maternal behavior and triggers local oxytocin release in lactating rats.Neuropharmacology. 2018 May 1;133:440-450. doi: 10.1016/j.neuropharm.2018.02.019. Epub 2018 Mar 2. Neuropharmacology. 2018. PMID: 29477300 Free PMC article.
-
When mothers neglect their offspring: an activated CRF system in the BNST is detrimental for maternal behavior.Arch Womens Ment Health. 2019 Jun;22(3):409-415. doi: 10.1007/s00737-018-0897-z. Epub 2018 Aug 4. Arch Womens Ment Health. 2019. PMID: 30078057 Review.
-
Mom doesn't care: When increased brain CRF system activity leads to maternal neglect in rodents.Front Neuroendocrinol. 2019 Apr;53:100735. doi: 10.1016/j.yfrne.2019.01.001. Epub 2019 Jan 23. Front Neuroendocrinol. 2019. PMID: 30684507 Review.
Cited by
-
Coordination of social behaviors by the bed nucleus of the stria terminalis.Eur J Neurosci. 2022 May;55(9-10):2404-2420. doi: 10.1111/ejn.14991. Epub 2020 Nov 7. Eur J Neurosci. 2022. PMID: 33006806 Free PMC article.
-
Maternally responsive neurons in the bed nucleus of the stria terminalis and medial preoptic area: Putative circuits for regulating anxiety and reward.Front Neuroendocrinol. 2015 Jul;38:65-72. doi: 10.1016/j.yfrne.2015.04.001. Epub 2015 Apr 21. Front Neuroendocrinol. 2015. PMID: 25910426 Free PMC article. Review.
-
Sex differences in fear responses: Neural circuits.Neuropharmacology. 2023 Jan 1;222:109298. doi: 10.1016/j.neuropharm.2022.109298. Epub 2022 Oct 31. Neuropharmacology. 2023. PMID: 36328063 Free PMC article. Review.
-
Corticotropin-releasing factor infusion in the bed nucleus of the stria terminalis of lactating mice alters maternal care and induces behavioural phenotypes in offspring.Sci Rep. 2020 Nov 17;10(1):19985. doi: 10.1038/s41598-020-77118-7. Sci Rep. 2020. PMID: 33204022 Free PMC article.
-
Corticotropin-releasing factor and GABA in the ventral tegmental area modulate partner preference formation in male and female prairie voles (Microtus ochrogaster).Front Neurosci. 2024 Jul 23;18:1430447. doi: 10.3389/fnins.2024.1430447. eCollection 2024. Front Neurosci. 2024. PMID: 39108311 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources