

Coupling mechanism and significance of the BOLD signal: a status report
- PMID: 25032494
- PMCID: PMC4147398
- DOI: 10.1146/annurev-neuro-071013-014111
Coupling mechanism and significance of the BOLD signal: a status report
Abstract
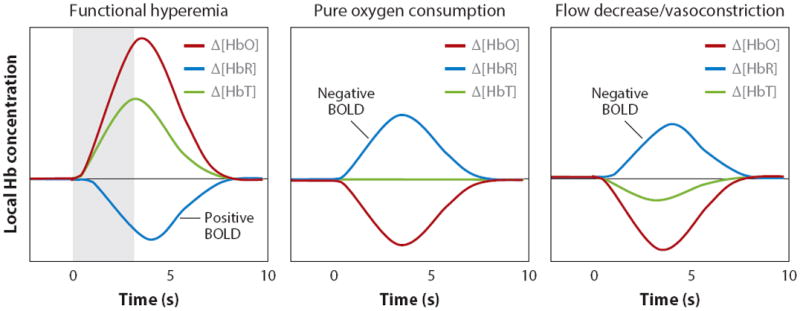
Functional magnetic resonance imaging (fMRI) provides a unique view of the working human mind. The blood-oxygen-level-dependent (BOLD) signal, detected in fMRI, reflects changes in deoxyhemoglobin driven by localized changes in brain blood flow and blood oxygenation, which are coupled to underlying neuronal activity by a process termed neurovascular coupling. Over the past 10 years, a range of cellular mechanisms, including astrocytes, pericytes, and interneurons, have been proposed to play a role in functional neurovascular coupling. However, the field remains conflicted over the relative importance of each process, while key spatiotemporal features of BOLD response remain unexplained. Here, we review current candidate neurovascular coupling mechanisms and propose that previously overlooked involvement of the vascular endothelium may provide a more complete picture of how blood flow is controlled in the brain. We also explore the possibility and consequences of conditions in which neurovascular coupling may be altered, including during postnatal development, pathological states, and aging, noting relevance to both stimulus-evoked and resting-state fMRI studies.
Keywords: astrocytes; fMRI; neurovascular coupling; pericytes; vascular endothelium.
Figures
References
-
- Ances BM, Buerk DG, Greenberg JH, Detre JA. Temporal dynamics of the partial pressure of brain tissue oxygen during functional forepaw stimulation in rats. Neurosci Lett. 2001;306:106–10. - PubMed
-
- Andresen J, Shafi NI, Bryan RM., Jr Endothelial influences on cerebrovascular tone. J Appl Physiol. 2006;100:318–27. - PubMed
-
- Armulik A, Abramsson A, Betsholtz C. Endothelial/pericyte interactions. Circ Res. 2005;97:512–23. - PubMed
-
- Arnerić SP, Honig MA, Milner TA, Greco S, Iadecola C, Reis DJ. Neuronal and endothelial sites of acetylcholine synthesis and release associated with microvessels in rat cerebral cortex: ultrastructural and neurochemical studies. Brain Res. 1988;454:11–30. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
