

Larger mammalian body size leads to lower retroviral activity
- PMID: 25033295
- PMCID: PMC4102558
- DOI: 10.1371/journal.ppat.1004214
Larger mammalian body size leads to lower retroviral activity
Abstract
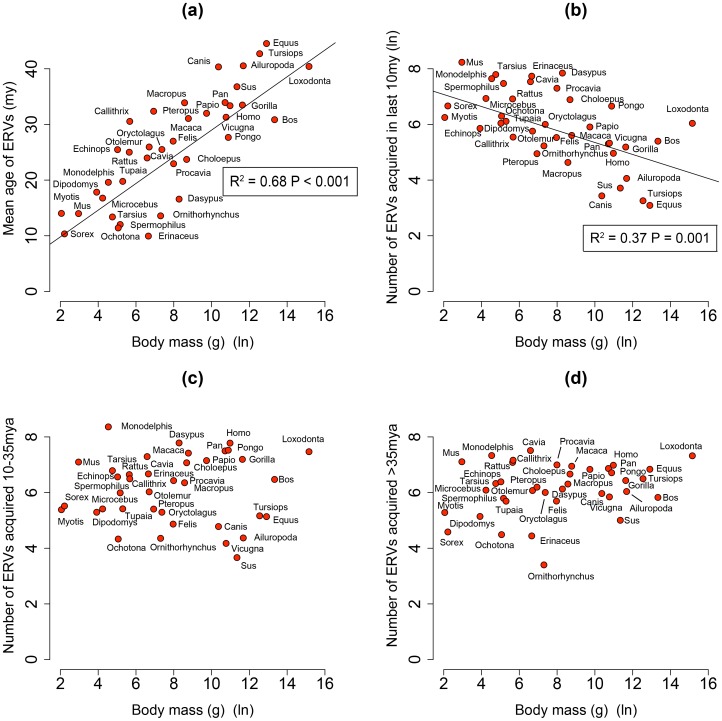
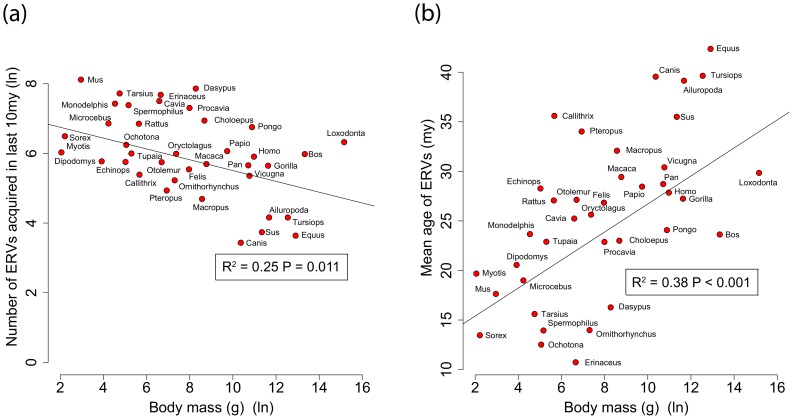
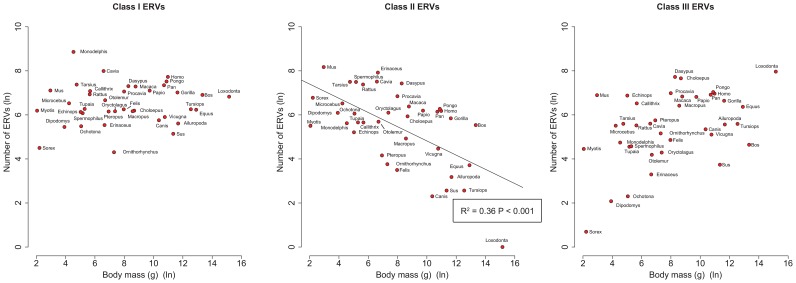
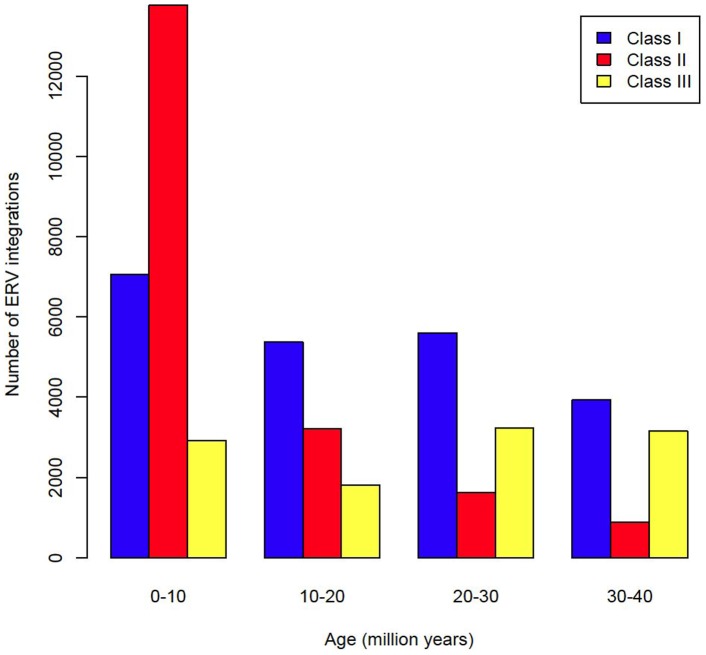
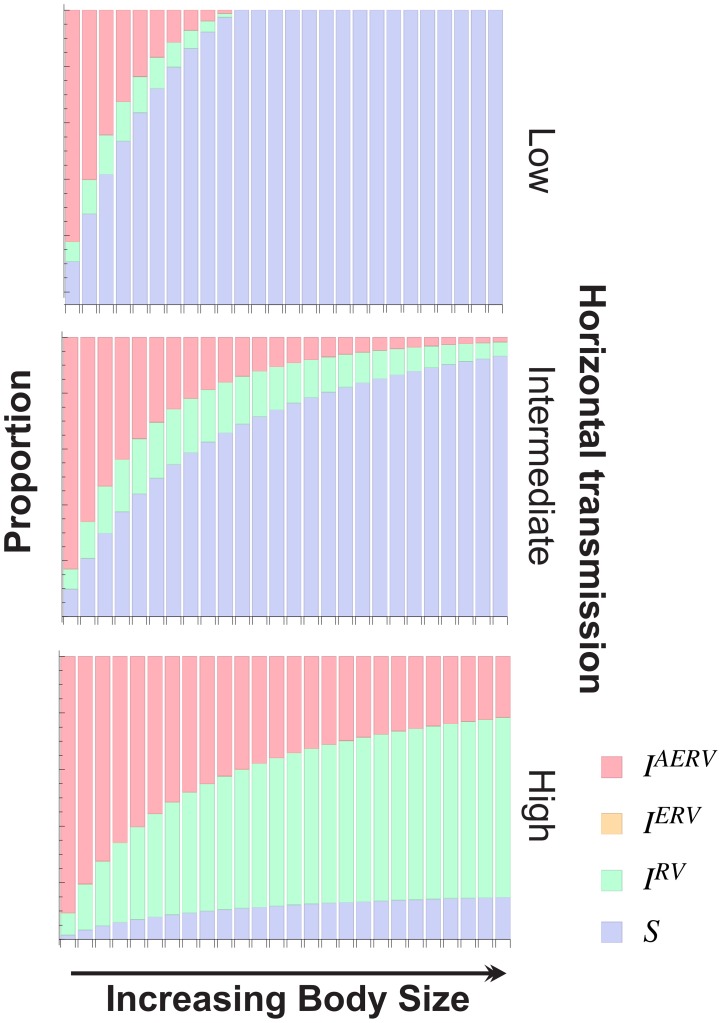
Retroviruses have been infecting mammals for at least 100 million years, leaving descendants in host genomes known as endogenous retroviruses (ERVs). The abundance of ERVs is partly determined by their mode of replication, but it has also been suggested that host life history traits could enhance or suppress their activity. We show that larger bodied species have lower levels of ERV activity by reconstructing the rate of ERV integration across 38 mammalian species. Body size explains 37% of the variance in ERV integration rate over the last 10 million years, controlling for the effect of confounding due to other life history traits. Furthermore, 68% of the variance in the mean age of ERVs per genome can also be explained by body size. These results indicate that body size limits the number of recently replicating ERVs due to their detrimental effects on their host. To comprehend the possible mechanistic links between body size and ERV integration we built a mathematical model, which shows that ERV abundance is favored by lower body size and higher horizontal transmission rates. We argue that because retroviral integration is tumorigenic, the negative correlation between body size and ERV numbers results from the necessity to reduce the risk of cancer, under the assumption that this risk scales positively with body size. Our model also fits the empirical observation that the lifetime risk of cancer is relatively invariant among mammals regardless of their body size, known as Peto's paradox, and indicates that larger bodied mammals may have evolved mechanisms to limit ERV activity.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Bannert N, Kurth R (2006) The evolutionary dynamics of human endogenous retroviral families. Annu Rev Genomics Hum Genet 7: 149–173. - PubMed
-
- Katzourakis A, Tristem M (2005) Phylogeny of human endogenous and exogenous retroviruses. In: Sverdlov ED, editor. Retroviruses and primate genome evolution. Austin, TX: Landes Bioscience. pp. 186–203.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
