

WRKY45-dependent priming of diterpenoid phytoalexin biosynthesis in rice and the role of cytokinin in triggering the reaction
- PMID: 25033935
- PMCID: PMC4133022
- DOI: 10.1007/s11103-014-0221-x
WRKY45-dependent priming of diterpenoid phytoalexin biosynthesis in rice and the role of cytokinin in triggering the reaction
Abstract
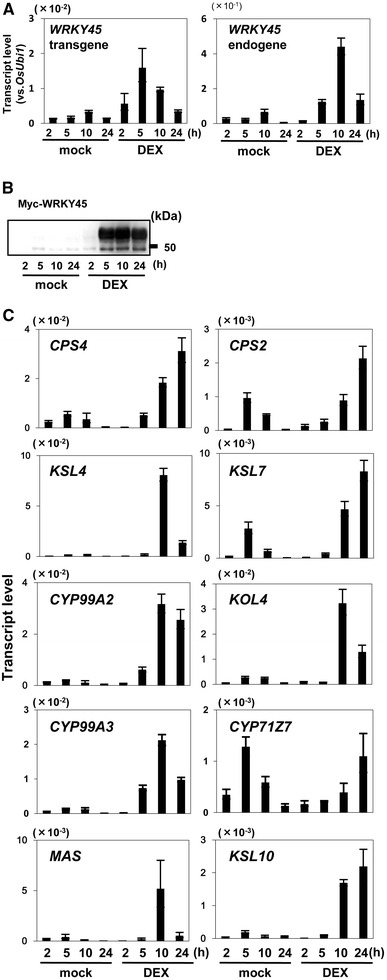
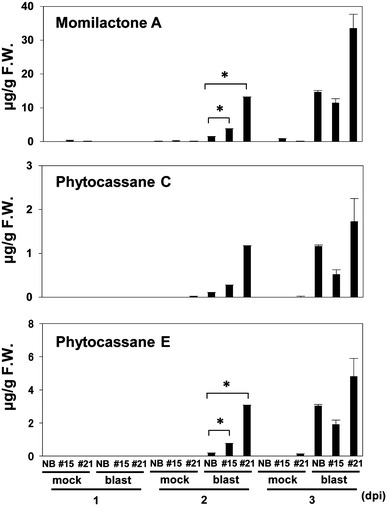
Plant activators such as benzothiadiazole (BTH) protect plants against diseases by priming the salicylic acid (SA) signaling pathway. In rice, the transcription factor WRKY45 plays a central role in this process. To investigate the mechanism involved in defense-priming by BTH and the role of WRKY45 in this process, we analyzed the transcripts of biosynthetic genes for diterpenoid phytoalexins (DPs) during the rice-Magnaporthe oryzae interaction. The DP biosynthetic genes were barely upregulated in BTH-treated rice plants, but were induced rapidly after M. oryzae infection in a WRKY45-dependent manner. These results indicate that the DP biosynthetic genes were primed by BTH through WRKY45. Rapid induction of the DP biosynthetic genes was also observed after M. oryzae infection to WRKY45-overexpressing (WRKY45-ox) plants. The changes in gene transcription resulted in accumulation of DPs in WRKY45-ox and BTH-pretreated rice after M. oryzae infection. Previously, we reported that cytokinins (CKs), especially isopentenyladenines, accumulated in M. oryzae-infected rice. Here, we show that DP biosynthetic genes are regulated by the SA/CK synergism in a WRKY45-dependent manner. Together, we propose that CK plays a role in mediating the signal of M. oryzae infection to trigger the induction of DP biosynthetic genes in BTH-primed plants.
Figures






References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
