

Density related effects on lifetime fecundity of Heterakis gallinarum in chickens
- PMID: 25034471
- PMCID: PMC4223411
- DOI: 10.1186/1756-3305-7-334
Density related effects on lifetime fecundity of Heterakis gallinarum in chickens
Abstract
Background: Density related effects, both inverse density- and density-dependent, contribute to regulating population dynamics of parasites. We investigated whether density related effects are directly controlling lifetime fecundity of Heterakis gallinarum.
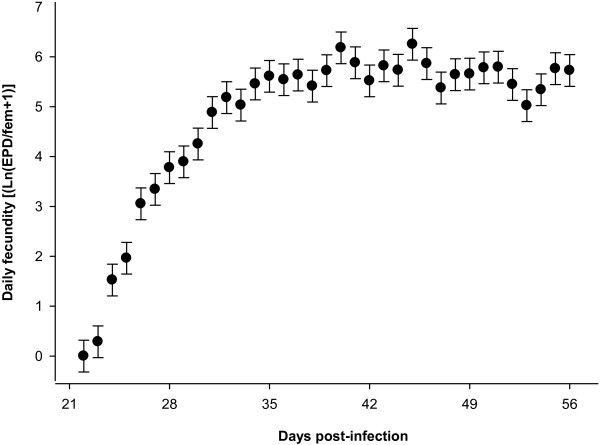
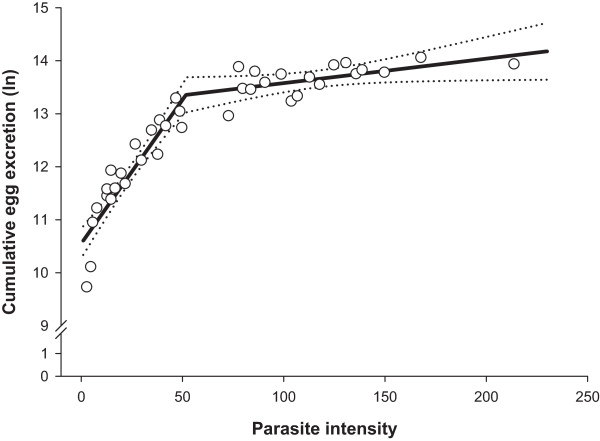
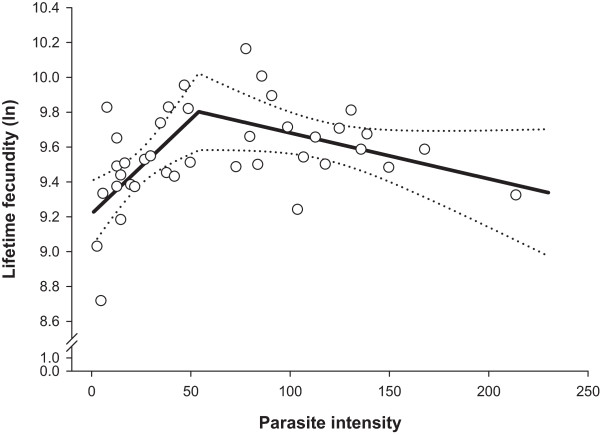
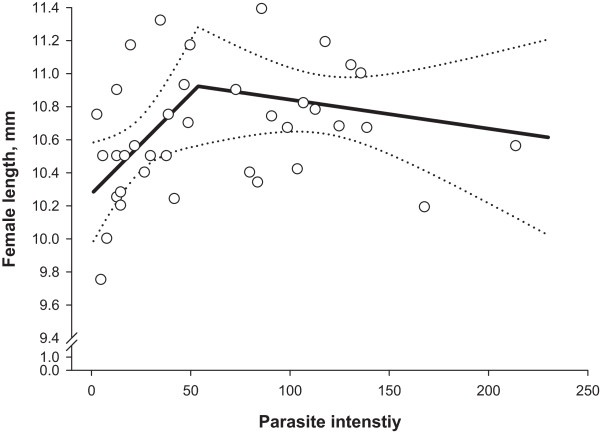
Methods: Daily total numbers of H. gallinarum eggs in faeces samples (N = 1365) from chickens (N = 39) were quantified starting from 3 weeks (wk) post-infection (p.i.). The birds were necropsied 8 wk p.i., and intensity and demographic characteristics of infrapopulations were determined. Density related effects on cumulative egg excretion (CEE), lifetime fecundity and worm length were investigated with a segmented regression analysis.
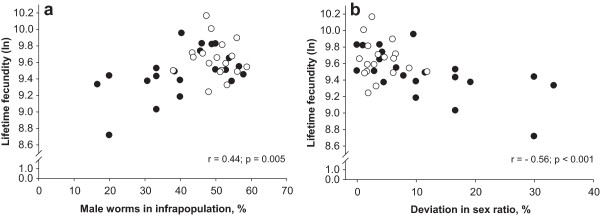
Results: For CEE, lifetime fecundity and female worm length, we determined highly similar parasite intensity thresholds (52-54 worms), which separated infrapopulations for influences of inverse density- and density dependence. CEE increased as parasite intensity increased up to an intensity of 52 worms. After this threshold, the relationship followed more of a horizontal line indicating impaired worm fecundity at higher parasite intensities. Lifetime fecundity was enhanced linearly in infrapopulations with up to 54 worms, but thereafter decreased gradually with increasing infrapopulation size. Female worm length increased linearly with elevating parasite intensity up to a threshold of 54 worms and thereafter declined with a rate of -0.014 mm for each additional worm. Lifetime fecundity and female worm length did not significantly differ between infrapopulations below and above the thresholds (P > 0.05). Lifetime fecundity was positively associated with the percentage of male worms (r = 0.44; P < 0.001), but negatively with absolute deviation from the theoretically expected sex-ratio in the infrapopulations (r = -0.56; P = 0.005). These relationships were stronger in infrapopulations below the threshold (r = 0.51 and -0.61, respectively), and were not significantly different from zero in the infrapopulations above the threshold (P > 0.05).
Conclusions: Egg production of H. gallinarum is regulated by the effects of both inverse density- and density-dependent mechanisms, which result in similar average lifetime fecundity below or above intensity thresholds. In infrapopulations below the intensity thresholds, inverse density dependence effects on lifetime fecundity appear to result partly from sex-ratio fluctuations and impaired mating success of the nematode.
Figures
References
-
- Keymer A. Density-dependent mechanisms in the regulation of intestinal helminth populations. Parasitology. 1982;84:573–587. - PubMed
-
- Stear MJ, Strain S, Bishop SC. Mechanisms underlying resistance to nematode infection. Int J Parasitol. 1999;29:51–56. - PubMed
-
- Paterson S, Viney ME. Host immune responses are necessary for density dependence in nematode infections. Parasitology. 2002;125:283–292. - PubMed
-
- Permin A, Bojesen M, Nansen P, Bisgaard M, Frandsen F, Pearman M. Ascaridia galli populations in chickens following single infections with different dose levels. Parasitol Res. 1997;83:614–617. - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
