

Structural insights into the low pH adaptation of a unique carboxylesterase from Ferroplasma: altering the pH optima of two carboxylesterases
- PMID: 25043762
- PMCID: PMC4148875
- DOI: 10.1074/jbc.M113.521856
Structural insights into the low pH adaptation of a unique carboxylesterase from Ferroplasma: altering the pH optima of two carboxylesterases
Abstract
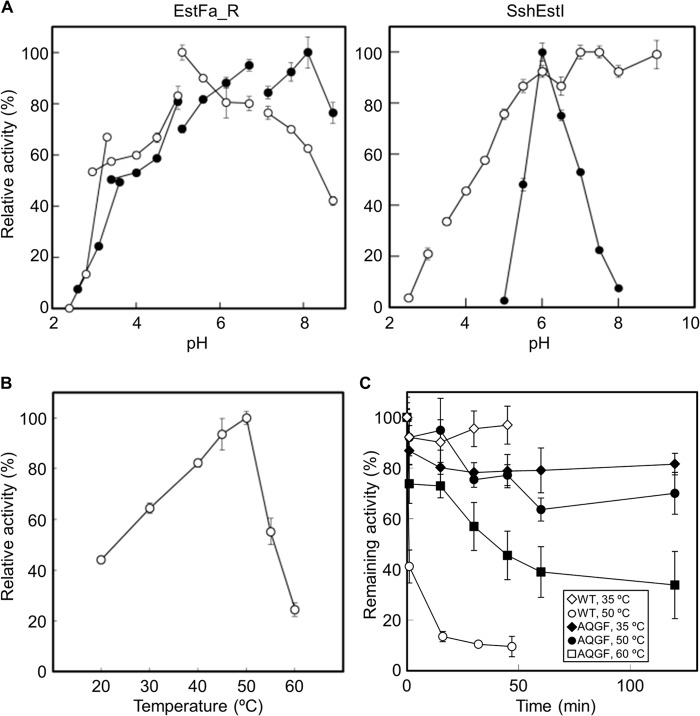
To investigate the mechanism for low pH adaptation by a carboxylesterase, structural and biochemical analyses of EstFa_R (a recombinant, slightly acidophilic carboxylesterase from Ferroplasma acidiphilum) and SshEstI (an alkaliphilic carboxylesterase from Sulfolobus shibatae DSM5389) were performed. Although a previous proteomics study by another group showed that the enzyme purified from F. acidiphilum contained an iron atom, EstFa_R did not bind to iron as analyzed by inductively coupled plasma MS and isothermal titration calorimetry. The crystal structures of EstFa_R and SshEstI were determined at 1.6- and 1.5-Å resolutions, respectively. EstFa_R had a catalytic triad with an extended hydrogen bond network that was not observed in SshEstI. Quadruple mutants of both proteins were created to remove or introduce the extended hydrogen bond network. The mutation on EstFa_R enhanced its catalytic efficiency and gave it an alkaline pH optimum, whereas the mutation on SshEstI resulted in opposite effects (i.e. a decrease in the catalytic efficiency and a downward shift in the optimum pH). Our experimental results suggest that the low pH optimum of EstFa_R activity was a result of the unique extended hydrogen bond network in the catalytic triad and the highly negatively charged surface around the active site. The change in the pH optimum of EstFa_R happened simultaneously with a change in the catalytic efficiency, suggesting that the local flexibility of the active site in EstFa_R could be modified by quadruple mutation. These observations may provide a novel strategy to elucidate the low pH adaptation of serine hydrolases.
Keywords: Archaea; Carboxylesterase; Enzyme Mechanism; Enzyme Structure; Ferroplasma acidiphilum; Hydrolase; Serine Hydrolase; pH Adaptation.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures





References
-
- Jaeger K. E., Reetz M. T. (1998) Microbial lipases form versatile tools for biotechnology. Trends Biotechnol. 16, 396–403 - PubMed
-
- Jaeger K. E., Dijkstra B. W., Reetz M. T. (1999) Bacterial biocatalysts: molecular biology, three-dimensional structures, and biotechnological applications of lipases. Annu. Rev. Microbiol. 53, 315–351 - PubMed
-
- Wei Y., Contreras J. A., Sheffield P., Osterlund T., Derewenda U., Kneusel R. E., Matern U., Holm C., Derewenda Z. S. (1999) Crystal structure of brefeldin A esterase, a bacterial homolog of the mammalian hormone-sensitive lipase. Nat. Struct. Biol. 6, 340–345 - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
