

Kinetic and structural characterization of tunnel-perturbing mutants in Bradyrhizobium japonicum proline utilization A
- PMID: 25046425
- PMCID: PMC4131897
- DOI: 10.1021/bi5007404
Kinetic and structural characterization of tunnel-perturbing mutants in Bradyrhizobium japonicum proline utilization A
Abstract
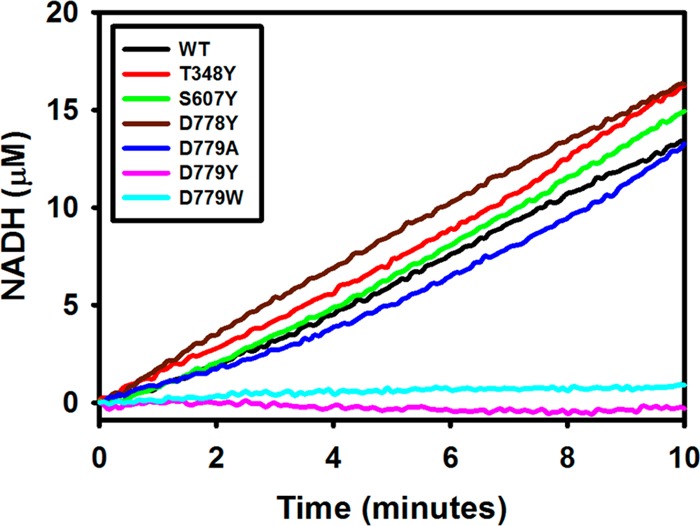
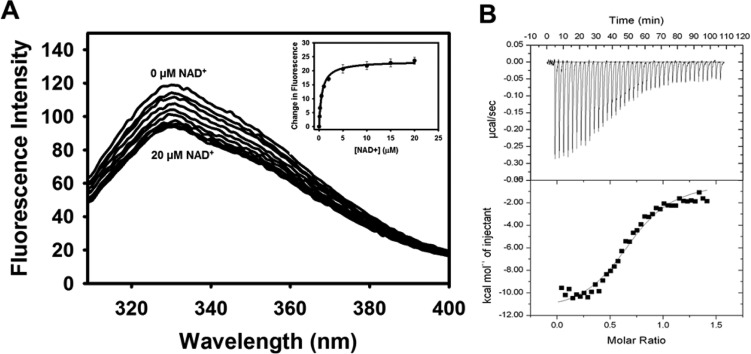
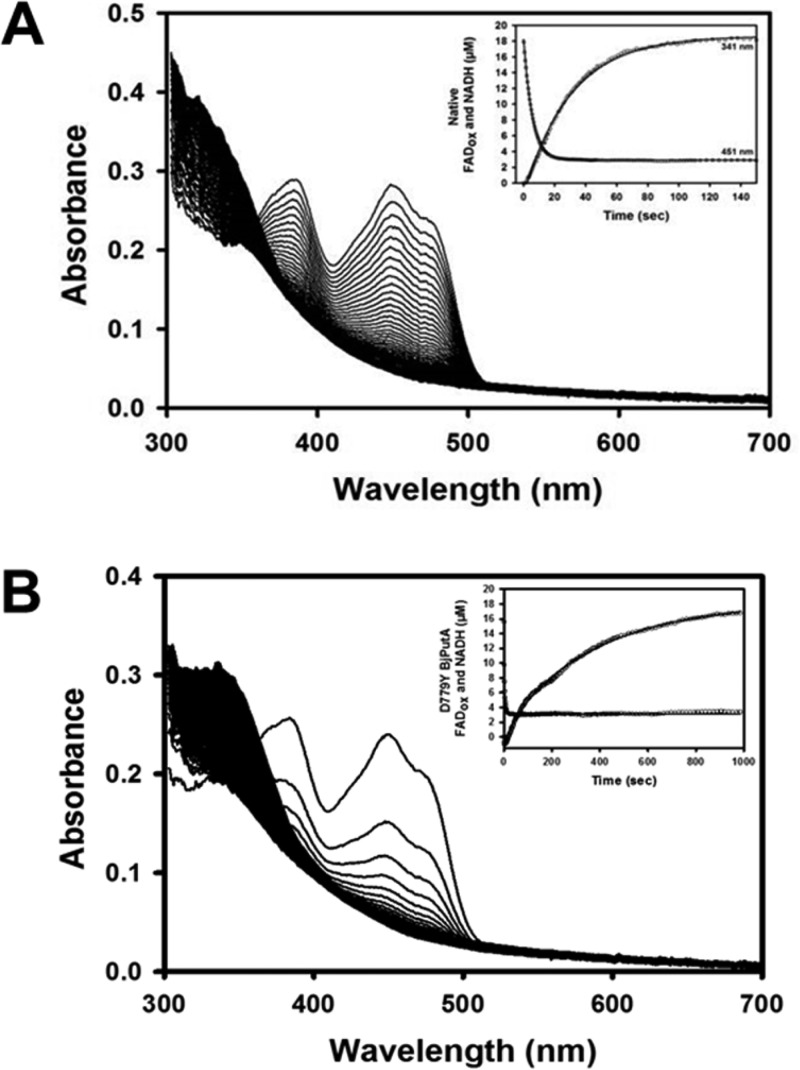
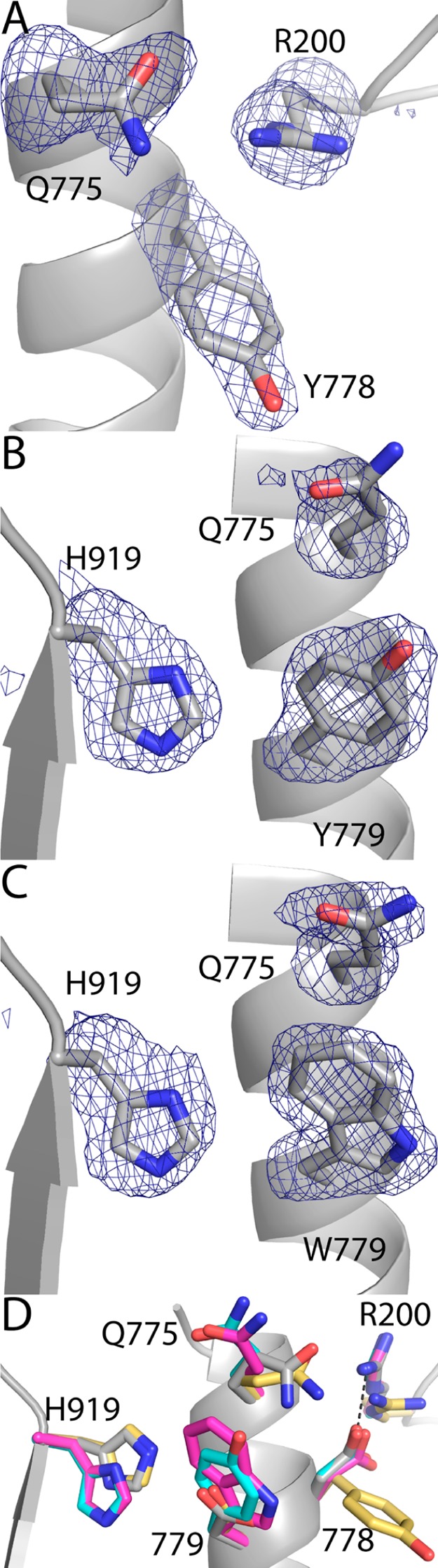
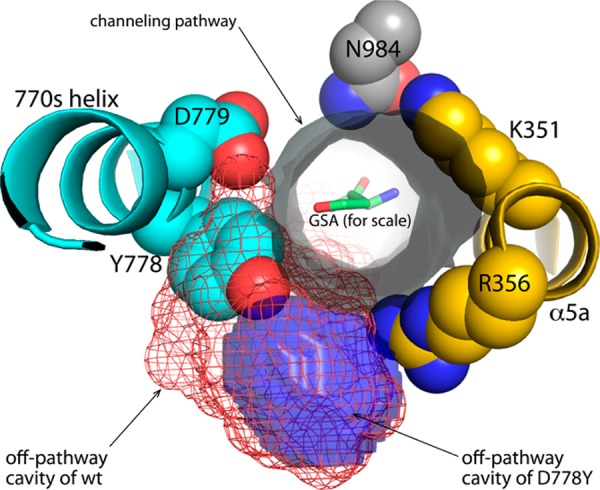
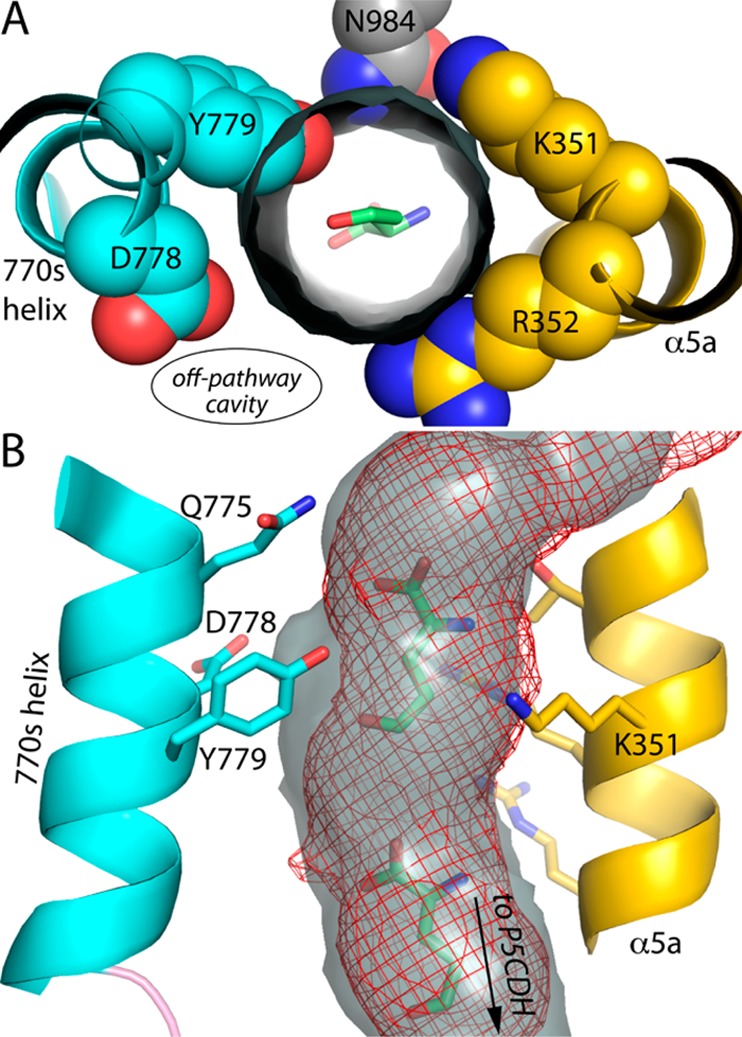
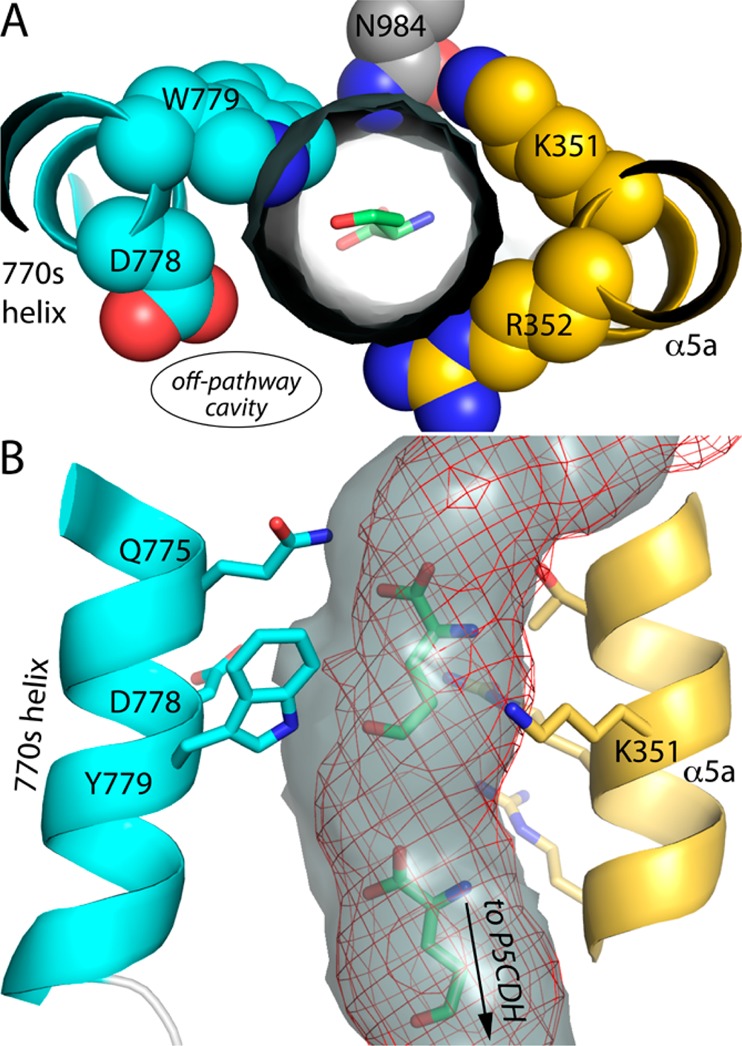
Proline utilization A from Bradyrhizobium japonicum (BjPutA) is a bifunctional flavoenzyme that catalyzes the oxidation of proline to glutamate using fused proline dehydrogenase (PRODH) and Δ(1)-pyrroline-5-carboxylate dehydrogenase (P5CDH) domains. Recent crystal structures and kinetic data suggest an intramolecular channel connects the two active sites, promoting substrate channeling of the intermediate Δ(1)-pyrroline-5-carboxylate/glutamate-γ-semialdehyde (P5C/GSA). In this work, the structure of the channel was explored by inserting large side chain residues at four positions along the channel in BjPutA. Kinetic analysis of the different mutants revealed replacement of D779 with Tyr (D779Y) or Trp (D779W) significantly decreased the overall rate of the PRODH-P5CDH channeling reaction. X-ray crystal structures of D779Y and D779W revealed that the large side chains caused a constriction in the central section of the tunnel, thus likely impeding the travel of P5C/GSA in the channel. The D779Y and D779W mutants have PRODH activity similar to that of wild-type BjPutA but exhibit significantly lower P5CDH activity, suggesting that exogenous P5C/GSA enters the channel upstream of Asp779. Replacement of nearby Asp778 with Tyr (D778Y) did not impact BjPutA channeling activity. Consistent with the kinetic results, the X-ray crystal structure of D778Y shows that the main channel pathway is not impacted; however, an off-cavity pathway is closed off from the channel. These findings provide evidence that the off-cavity pathway is not essential for substrate channeling in BjPutA.
Figures


References
-
- Nakajima K.; Inatsu S.; Mizote T.; Nagata Y.; Aoyama K.; Fukuda Y.; Nagata K. (2008) Possible involvement of putA gene in Helicobacter pylori colonization in the stomach and motility. Biomed. Res. 29, 9–18. - PubMed
-
- van Weelden S. W.; Fast B.; Vogt A.; van der Meer P.; Saas J.; van Hellemond J. J.; Tielens A. G.; Boshart M. (2003) Procyclic Trypanosoma brucei do not use Krebs cycle activity for energy generation. J. Biol. Chem. 278, 12854–12863. - PubMed
-
- Bringaud F.; Riviere L.; Coustou V. (2006) Energy metabolism of trypanosomatids: Adaptation to available carbon sources. Mol. Biochem. Parasitol. 149, 1–9. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
