

Cellular mechanism of premature ventricular contraction-induced cardiomyopathy
- PMID: 25046857
- PMCID: PMC4252777
- DOI: 10.1016/j.hrthm.2014.07.022
Cellular mechanism of premature ventricular contraction-induced cardiomyopathy
Abstract
Background: Frequent premature ventricular contractions (PVCs) are associated with increased risk of sudden cardiac death and can cause secondary cardiomyopathy.
Objective: We sought to determine the mechanism(s) responsible for prolonged refractory period and left ventricular (LV) dysfunction demonstrated in our canine model of PVC-induced cardiomyopathy.
Methods: Single myocytes were isolated from LV free wall of PVC and control canines and used for patch-clamp recording, intracellular Ca(2+) measurements, and immunocytochemistry/confocal microscopy. LV tissues adjacent to the area of myocyte isolation were used for the immunoblot quantification of protein expression.
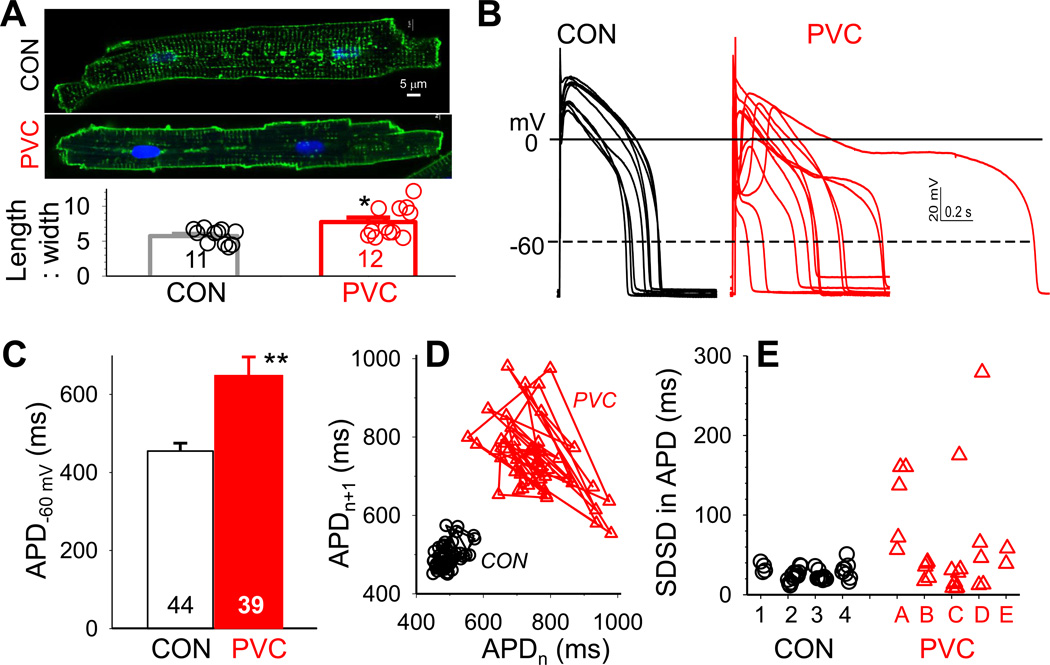
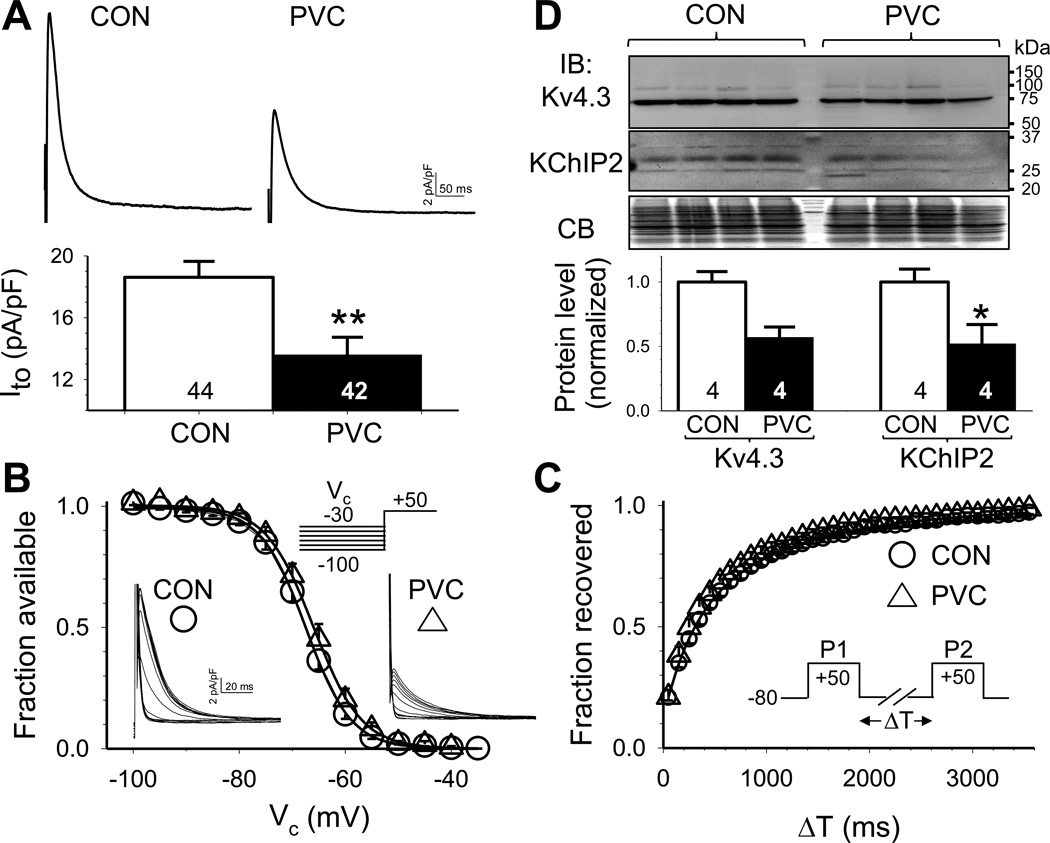
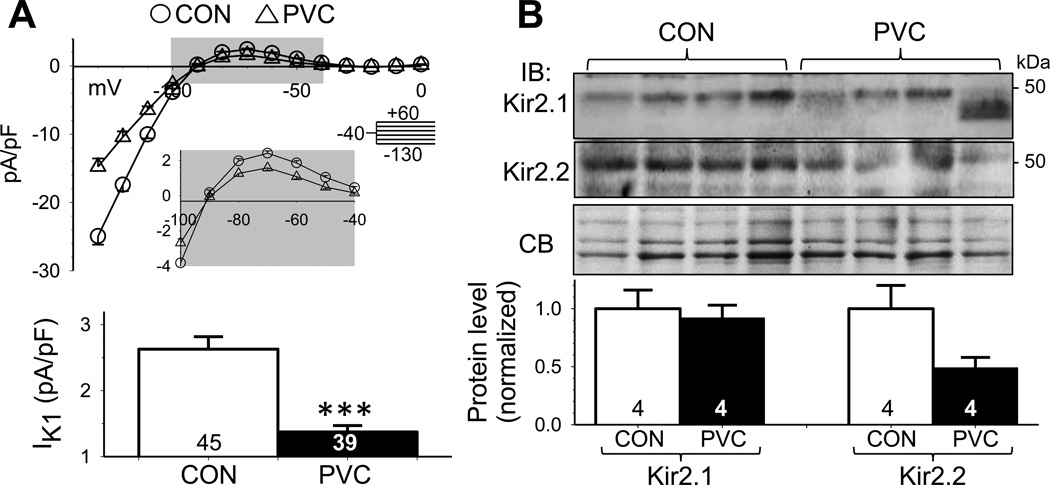
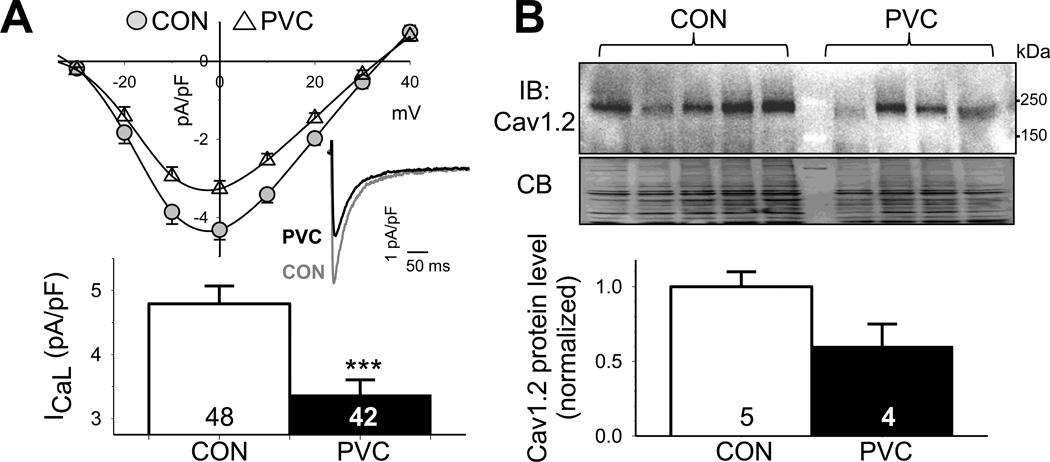
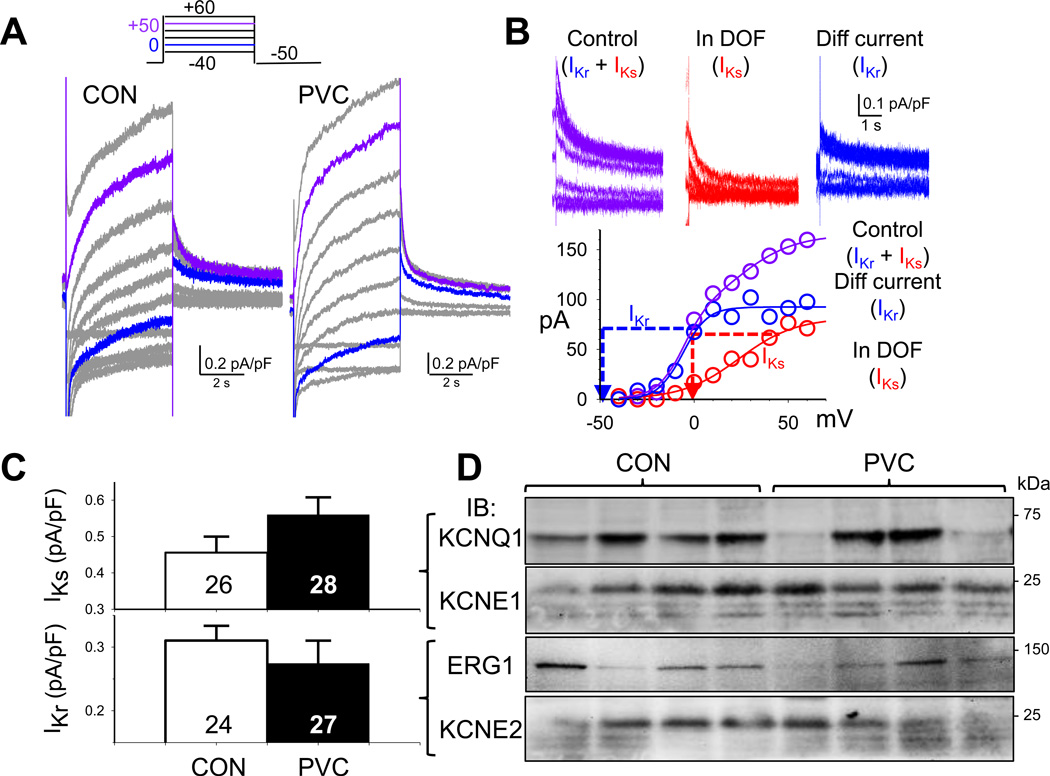
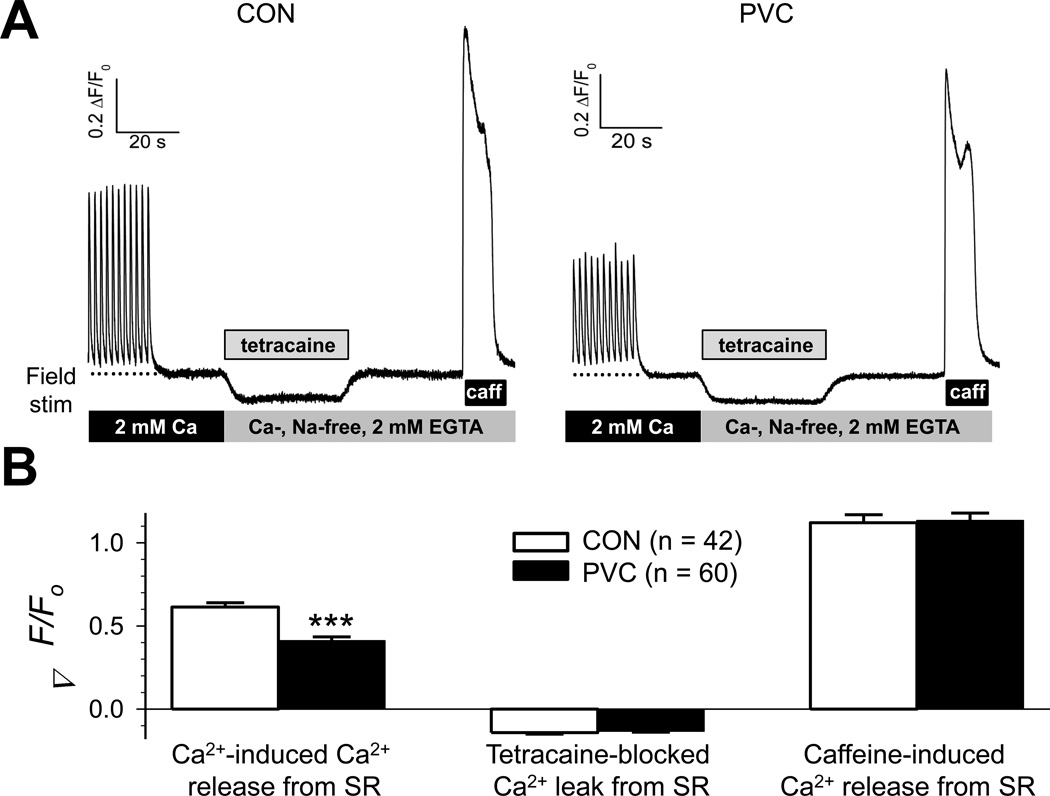
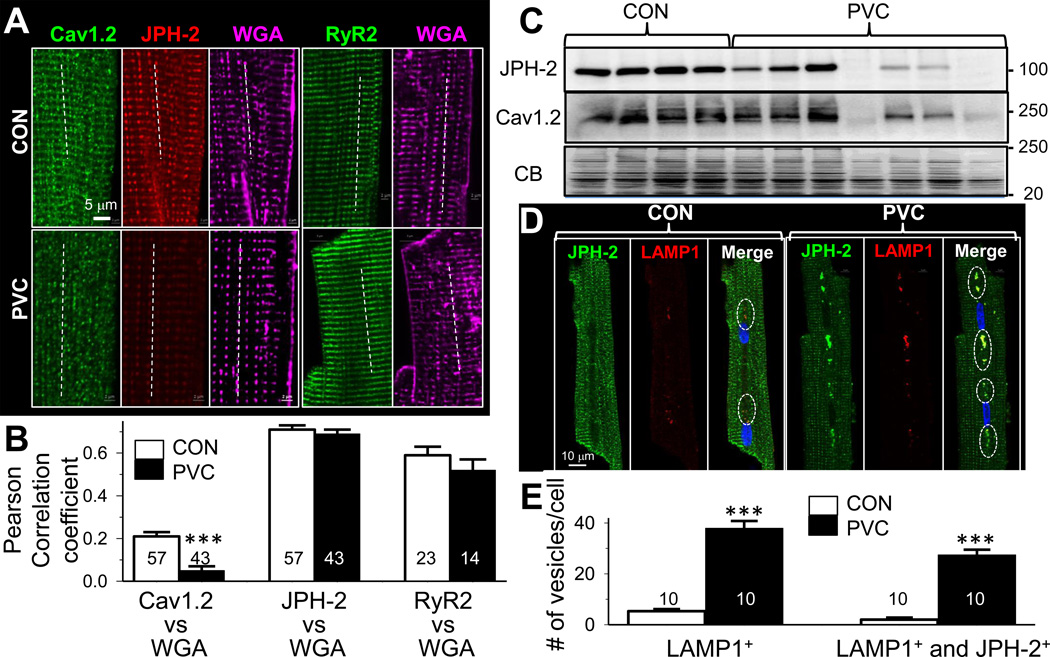
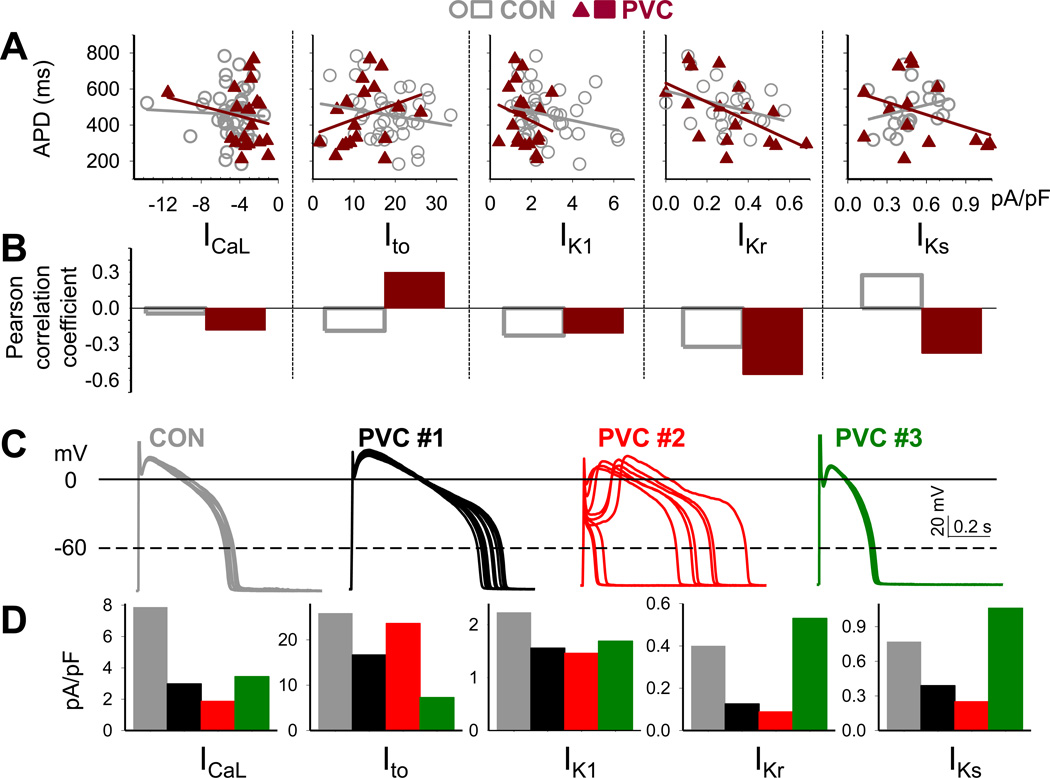
Results: In the PVC group, LV ejection fraction decreased from 57.6% ± 1.5% to 30.4% ± 3.1% after ≥4 months of ventricular bigeminy. Compared to control myocytes, PVC myocytes had decreased densities of both outward (transient outward current [Ito] and inward rectifier current [IK1]) and inward (L-type Ca current [ICaL]) currents, but no consistent changes in rapid or slow delayed rectifier currents. The reduction in Ito, IK1, and ICaL was accompanied by decreased protein levels of their channel subunits. The extent of reduction in Ito, IK1, and ICaL varied among PVC myocytes, creating marked heterogeneity in action potential configurations and durations. PVC myocytes showed impaired Ca-induced Ca release from the sarcoplasmic reticulum (SR), without increase in SR Ca leak or decrease in SR Ca store. This was accompanied by a decrease in dyad scaffolding protein, junctophilin-2, and loss of Cav1.2 registry with Ca-releasing channels (ryanodine receptor 2).
Conclusion: PVCs increase dispersion of action potential configuration/duration, a risk factor for sudden cardiac death, because of the heterogeneous reduction in Ito, IK1, and ICaL. The excitation-contraction coupling is impaired because of the decrease in ICaL and Cav1.2 misalignment with respect to ryanodine receptor 2.
Keywords: Cardiomyopathy; Electrical remodeling; Excitation-contraction coupling; Premature ventricular contraction.
Copyright © 2014 Heart Rhythm Society. All rights reserved.
Figures
Comment in
-
The consequences of contracting early and often.Heart Rhythm. 2014 Nov;11(11):2073-4. doi: 10.1016/j.hrthm.2014.08.015. Epub 2014 Aug 12. Heart Rhythm. 2014. PMID: 25131179 No abstract available.
References
-
- Liu X-S, Jiang M, Zhang M, Tang D, Higgins RSD, Tseng G-N. Electrical remodeling in a canine model of ischemic cardiomyopathy. Am J Physiol. 2007;292:H560–H571. - PubMed
-
- Brennan M, Palaniswami M, Kamen P. Do existing measures of Poincare plot geometry reflect nonlinear features of heart rate variability? IEEE Transactions on Biomedical Engineering. 2001;48:1342–1347. - PubMed
-
- Greenstein JL, Wu R, Po S, Tomaselli GF, Winslow RL. Role of the calcium-independent transient outward current Ito1 in shaping action potential morphology and duration. Circ Res. 2000;87:1026–1033. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
