

Pericentriolar material structure and dynamics
- PMID: 25047613
- PMCID: PMC4113103
- DOI: 10.1098/rstb.2013.0459
Pericentriolar material structure and dynamics
Abstract
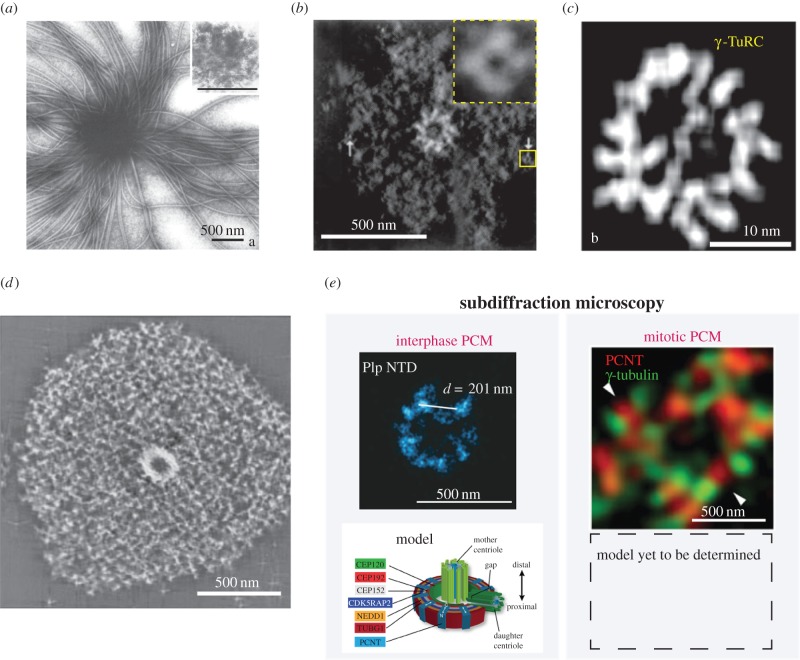
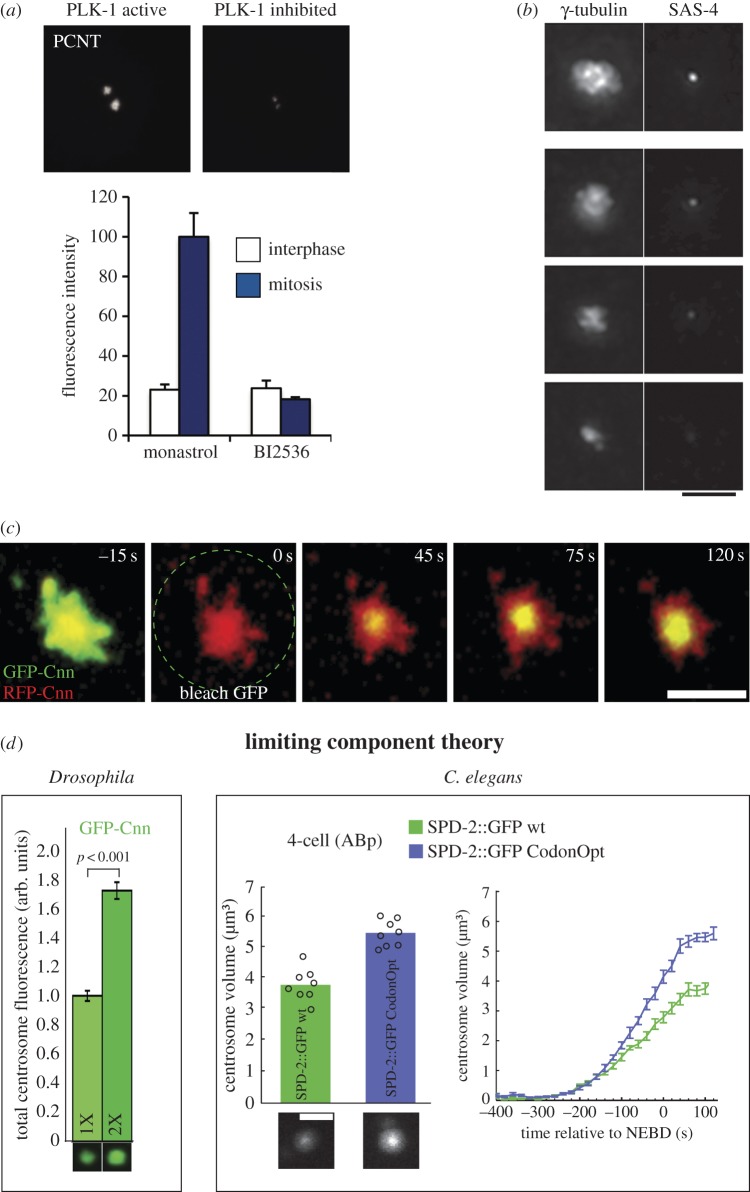
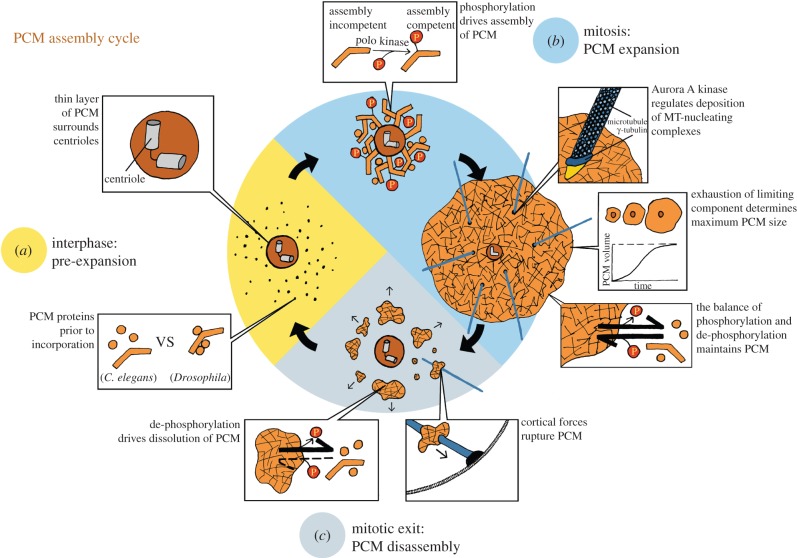
A centrosome consists of two barrel-shaped centrioles embedded in a matrix of proteins known as the pericentriolar material (PCM). The PCM serves as a platform for protein complexes that regulate organelle trafficking, protein degradation and spindle assembly. Perhaps most important for cell division, the PCM concentrates tubulin and serves as the primary organizing centre for microtubules in metazoan somatic cells. Thus, similar to other well-described organelles, such as the nucleus and mitochondria, the cell has compartmentalized a multitude of vital biochemical reactions in the PCM. However, unlike these other organelles, the PCM is not membrane bound, but rather a dynamic collection of protein complexes and nucleic acids that constitute the organelle's interior and determine its boundary. How is the complex biochemical machinery necessary for the myriad centrosome functions concentrated and maintained in the PCM? Recent advances in proteomics and RNAi screening have unveiled most of the key PCM components and hinted at their molecular interactions ( table 1). Now we must understand how the interactions between these molecules contribute to the mesoscale organization and the assembly of the centrosome. Among outstanding questions are the intrinsic mechanisms that determine PCM shape and size, and how it functions as a biochemical reaction hub.
Keywords: centrosome; microtubule-organizing centre; organelle scaling; pericentriolar material.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
