

The centriole duplication cycle
- PMID: 25047614
- PMCID: PMC4113104
- DOI: 10.1098/rstb.2013.0460
The centriole duplication cycle
Abstract
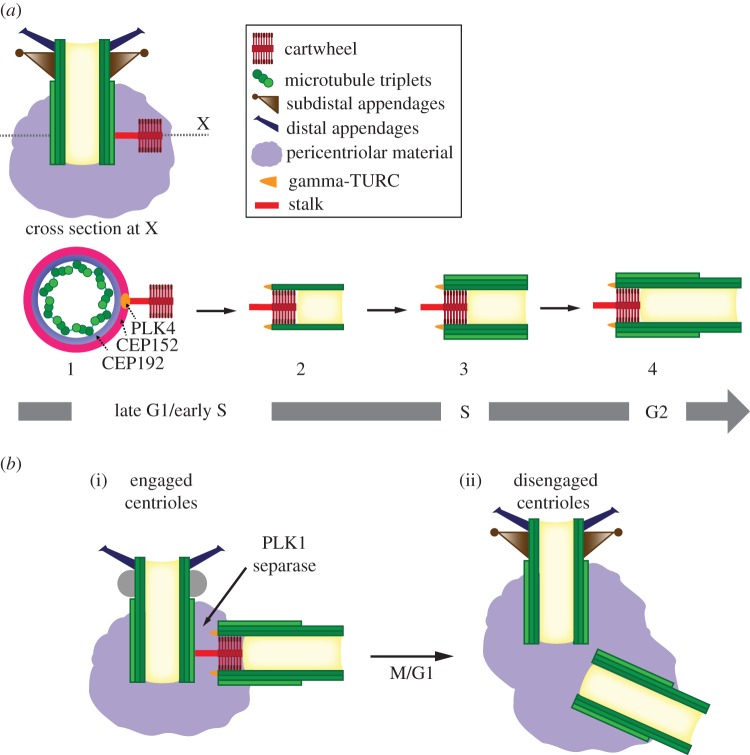
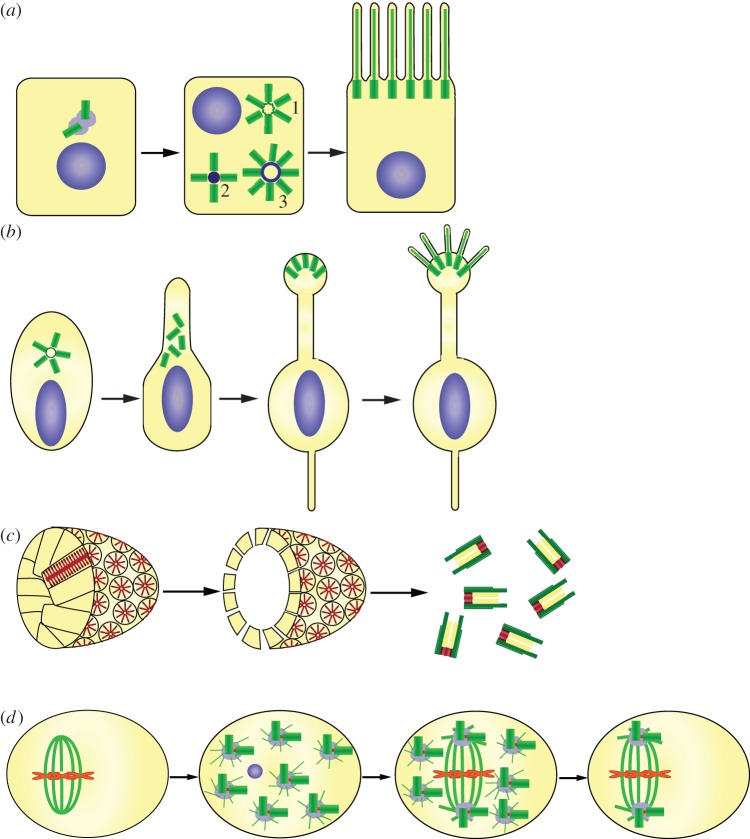
Centrosomes are the main microtubule-organizing centre of animal cells and are important for many critical cellular and developmental processes from cell polarization to cell division. At the core of the centrosome are centrioles, which recruit pericentriolar material to form the centrosome and act as basal bodies to nucleate formation of cilia and flagella. Defects in centriole structure, function and number are associated with a variety of human diseases, including cancer, brain diseases and ciliopathies. In this review, we discuss recent advances in our understanding of how new centrioles are assembled and how centriole number is controlled. We propose a general model for centriole duplication control in which cooperative binding of duplication factors defines a centriole 'origin of duplication' that initiates duplication, and passage through mitosis effects changes that license the centriole for a new round of duplication in the next cell cycle. We also focus on variations on the general theme in which many centrioles are created in a single cell cycle, including the specialized structures associated with these variations, the deuterosome in animal cells and the blepharoplast in lower plant cells.
Keywords: blepharoplast; centriole; centrosome; deuterosome.
© 2014 The Author(s) Published by the Royal Society. All rights reserved.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
