

Heteromerization of GPR55 and cannabinoid CB2 receptors modulates signalling
- PMID: 25048571
- PMCID: PMC4294047
- DOI: 10.1111/bph.12850
Heteromerization of GPR55 and cannabinoid CB2 receptors modulates signalling
Abstract
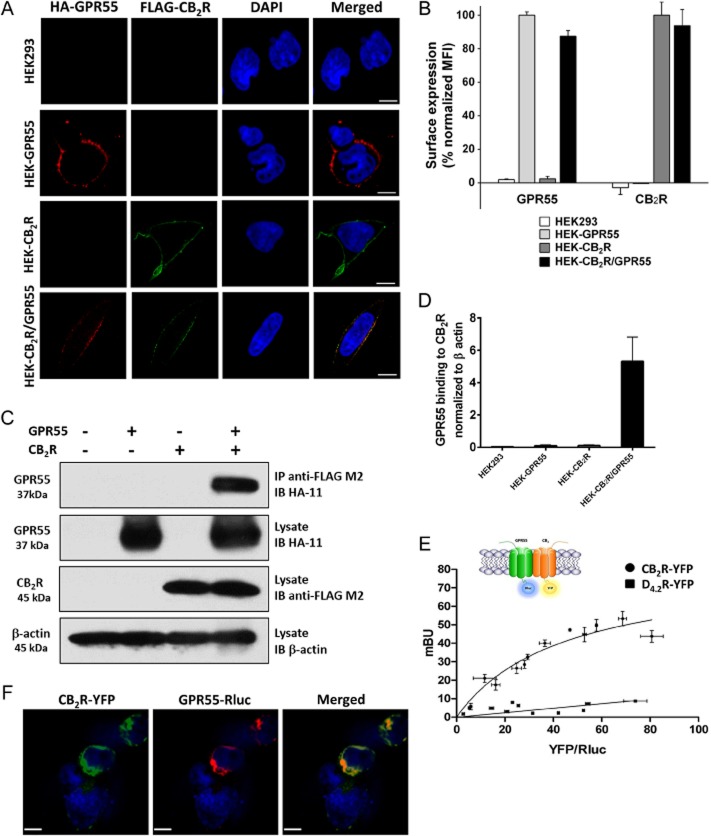
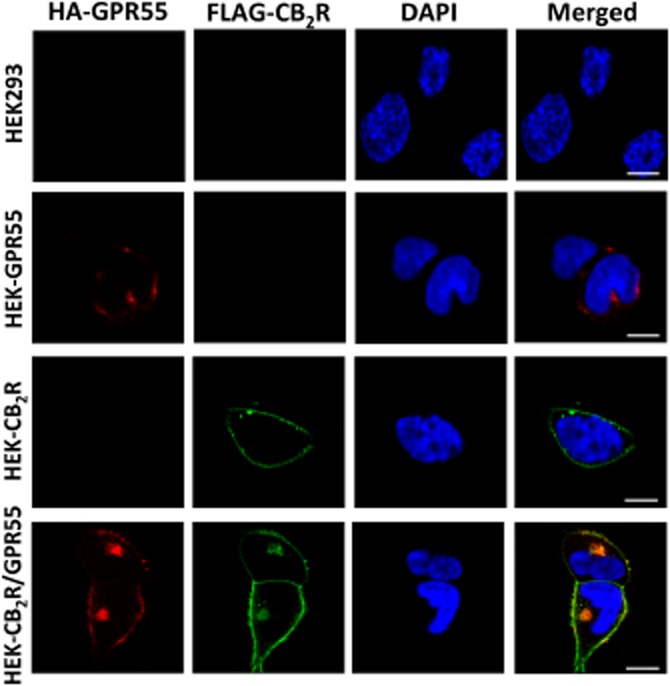
Background and purpose: Heteromerization of GPCRs is key to the integration of extracellular signals and the subsequent cell response via several mechanisms including heteromer-selective ligand binding, trafficking and/or downstream signalling. As the lysophosphatidylinositol GPCR 55 (GPR55) has been shown to affect the function of the cannabinoid receptor subtype 2 (CB2 receptor) in human neutrophils, we investigated the possible heteromerization of CB2 receptors with GPR55.
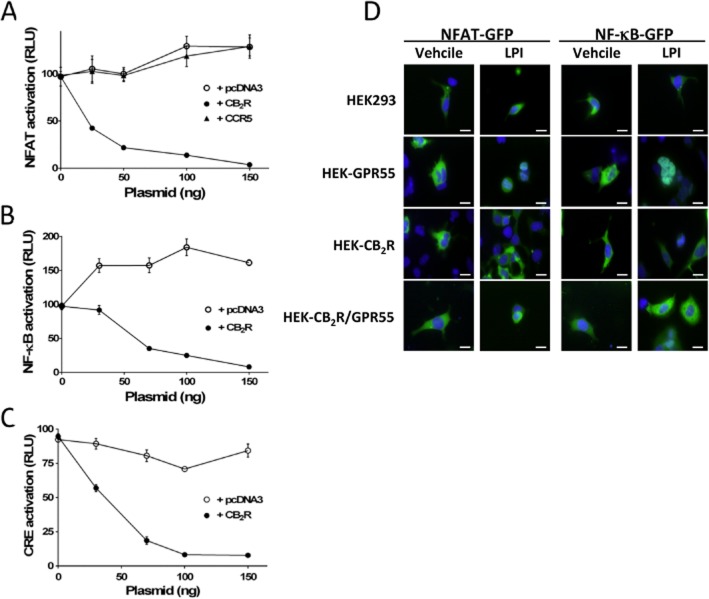
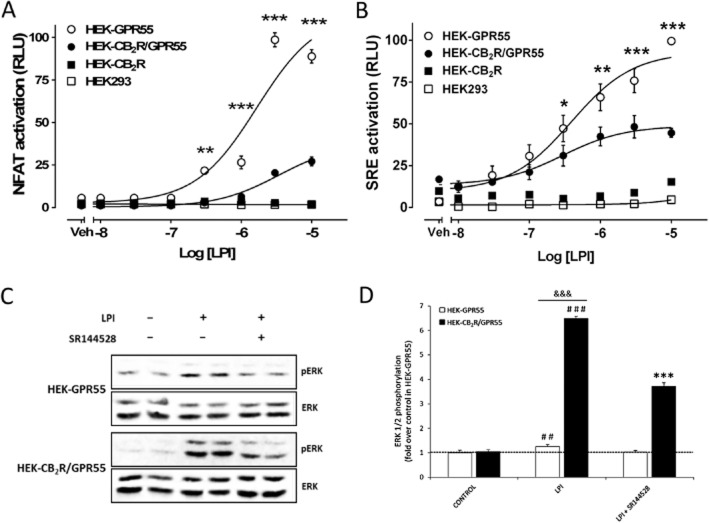
Experimental approach: The direct interaction of human GPR55 and CB2 receptors heterologously expressed in HEK293 cells was assessed by co-immunoprecipitation and bioluminescence resonance energy transfer assays. The effect of cross-talk on signalling was investigated at downstream levels by label-free real-time methods (Epic dynamic mass redistribution and CellKey impedance assays), ERK1/2-MAPK activation and gene reporter assays.
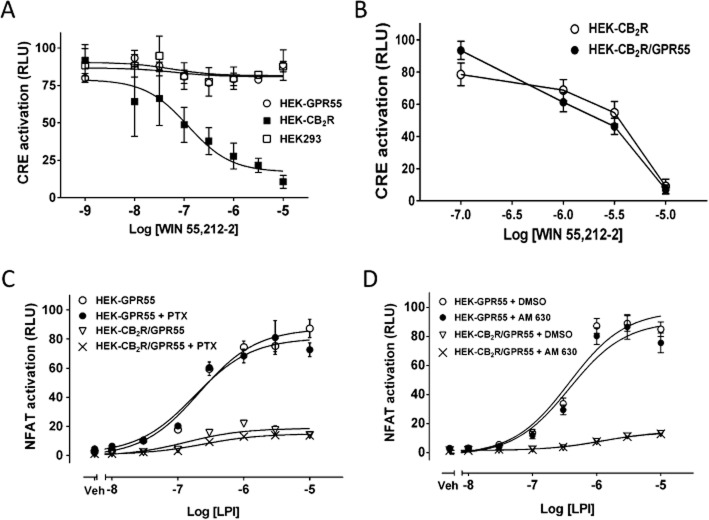
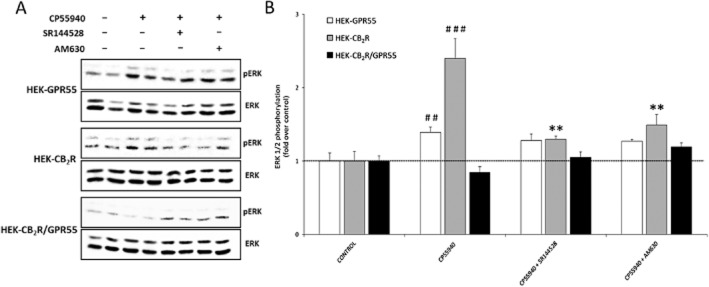
Key results: GPR55 and CB2 receptors co-localized on the surface of HEK293 cells, co-precipitated in membrane extracts and formed heteromers in living HEK293 cells. Whereas heteromerization led to a reduction in GPR55-mediated activation of transcription factors (nuclear factor of activated T-cells, NF-κB and cAMP response element), ERK1/2-MAPK activation was potentiated in the presence of CB2 receptors. CB2 receptor-mediated signalling was also affected by co-expression with GPR55. Label-free assays confirmed cross-talk between the two receptors.
Conclusions and implications: Heteromers, unique signalling units, form in HEK293 cells expressing GPR55 and CB2 receptors. The signalling by agonists of either receptor was governed (i) by the presence or absence of the partner receptors (with the consequent formation of heteromers) and (ii) by the activation state of the partner receptor.
© 2014 The British Pharmacological Society.
Figures




References
-
- Andradas C, Caffarel MM, Perez-Gomez E, Salazar M, Lorente M, Velasco G, et al. The orphan G protein-coupled receptor GPR55 promotes cancer cell proliferation via ERK. Oncogene. 2011;30:245–252. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
